

Hairpin dsRNA does not trigger RNA interference in Candida albicans cells
- PMID: 20737430
- PMCID: PMC4677786
- DOI: 10.1002/yea.1814
Hairpin dsRNA does not trigger RNA interference in Candida albicans cells
Abstract
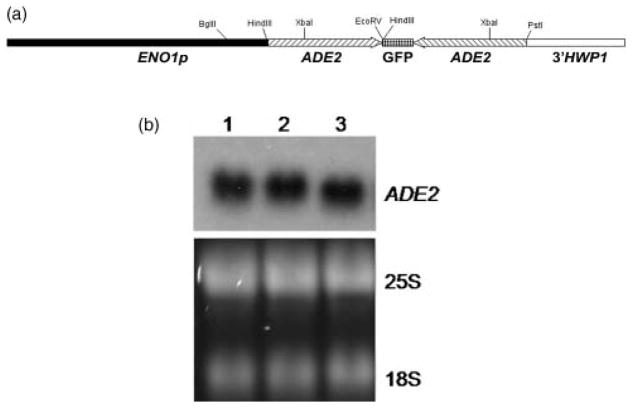
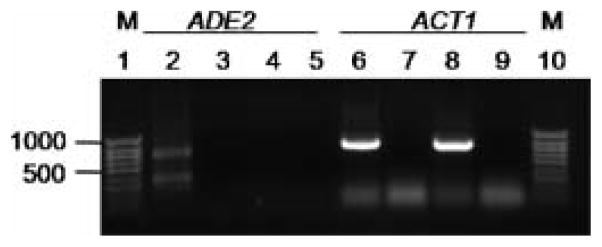
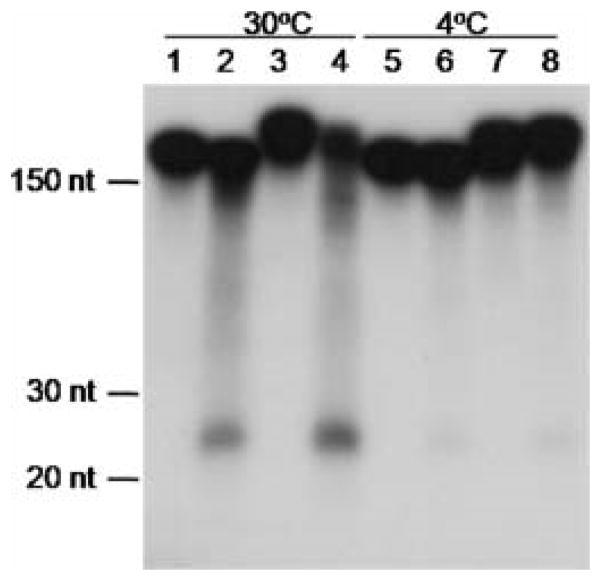
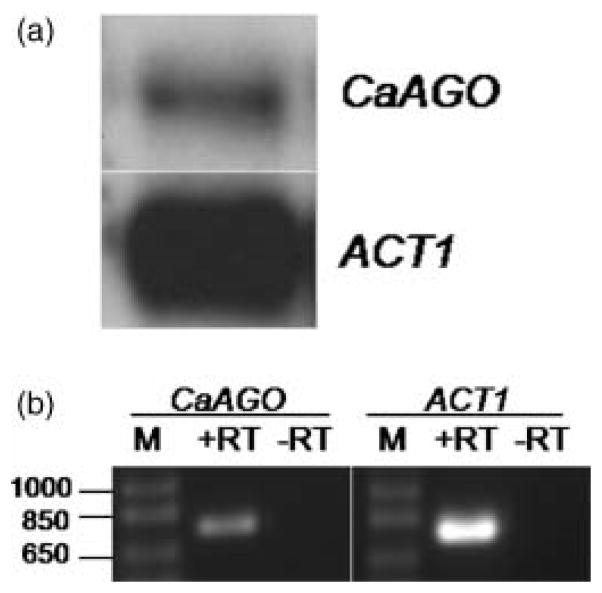
RNA interference/silencing mechanisms triggered by double-stranded RNA (dsRNA) have been described in many eukaryotes, including fungi. These mechanisms have in common small RNA molecules (siRNAs or microRNAs) originating from dsRNAs that, together with the effector protein Argonaute, mediate silencing. The genome of the fungal pathogen Candida albicans harbours a well-conserved Argonaute and a non-canonical Dicer, essential members of silencing pathways. Prototypical siRNAs are detected as members of the C. albicans transcriptome, which is potential evidence of RNA interference/silencing pathways in this organism. Surprisingly, expression of a dsRNA a hairpin ADE2 dsRNA molecule to interfere with the endogenous ADE2 mRNA did not result in down-regulation of the message or produce adenine auxotrophic strains. Cell free assays showed that the hairpin dsRNA was a substrate for the putative C. albicans Dicer, discounting the possibility that the nature of the dsRNA trigger affects silencing functionality. Our results suggested that unknown cellular events govern the functionality of siRNAs originating from transgenes in RNA interference/silencing pathways in C. albicans.
Copyright © 2010 John Wiley & Sons, Ltd.
Figures
Similar articles
-
Short hairpin type of dsRNAs that are controlled by tRNA(Val) promoter significantly induce RNAi-mediated gene silencing in the cytoplasm of human cells.Nucleic Acids Res. 2003 Jan 15;31(2):700-7. doi: 10.1093/nar/gkg158. Nucleic Acids Res. 2003. PMID: 12527779 Free PMC article.
-
Endogenous siRNAs from naturally formed dsRNAs regulate transcripts in mouse oocytes.Nature. 2008 May 22;453(7194):539-43. doi: 10.1038/nature06908. Epub 2008 Apr 10. Nature. 2008. PMID: 18404146
-
A double-stranded-RNA response program important for RNA interference efficiency.Mol Cell Biol. 2007 Jun;27(11):3995-4005. doi: 10.1128/MCB.00186-07. Epub 2007 Mar 19. Mol Cell Biol. 2007. PMID: 17371837 Free PMC article.
-
How does RNA editing affect dsRNA-mediated gene silencing?Cold Spring Harb Symp Quant Biol. 2006;71:285-92. doi: 10.1101/sqb.2006.71.037. Cold Spring Harb Symp Quant Biol. 2006. PMID: 17381308 Free PMC article. Review.
-
Plant dicer-like proteins: double-stranded RNA-cleaving enzymes for small RNA biogenesis.J Plant Res. 2017 Jan;130(1):33-44. doi: 10.1007/s10265-016-0877-1. Epub 2016 Nov 24. J Plant Res. 2017. PMID: 27885504 Review.
Cited by
-
Characterization of genetic determinants that modulate Candida albicans filamentation in the presence of bacteria.PLoS One. 2013 Aug 7;8(8):e71939. doi: 10.1371/journal.pone.0071939. eCollection 2013. PLoS One. 2013. PMID: 23951271 Free PMC article.
-
Milestones in Candida albicans gene manipulation.Fungal Genet Biol. 2011 Sep;48(9):858-65. doi: 10.1016/j.fgb.2011.04.003. Epub 2011 Apr 14. Fungal Genet Biol. 2011. PMID: 21511047 Free PMC article. Review.
-
RNA-mediated gene silencing in Candida albicans: inhibition of hyphae formation by use of RNAi technology.Mycopathologia. 2012 Sep;174(3):177-85. doi: 10.1007/s11046-012-9539-6. Epub 2012 Apr 7. Mycopathologia. 2012. PMID: 22484810
-
Down-regulation of the ALS3 gene as a consequent effect of RNA-mediated silencing of the EFG1 gene in Candida albicans.Iran Biomed J. 2012;16(4):172-8. doi: 10.6091/ibj.1093.2012. Iran Biomed J. 2012. PMID: 23183615 Free PMC article.
-
Novel antifungals and treatment approaches to tackle resistance and improve outcomes of invasive fungal disease.Clin Microbiol Rev. 2024 Jun 13;37(2):e0007423. doi: 10.1128/cmr.00074-23. Epub 2024 Apr 11. Clin Microbiol Rev. 2024. PMID: 38602408 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources