

The evolutionary history of histone H3 suggests a deep eukaryotic root of chromatin modifying mechanisms
- PMID: 20738881
- PMCID: PMC2939574
- DOI: 10.1186/1471-2148-10-259
The evolutionary history of histone H3 suggests a deep eukaryotic root of chromatin modifying mechanisms
Abstract
Background: The phenotype of an organism is an outcome of both its genotype, encoding the primary sequence of proteins, and the developmental orchestration of gene expression. The substrate of gene expression in eukaryotes is the chromatin, whose fundamental units are nucleosomes composed of DNA wrapped around each two of the core histone types H2A, H2B, H3 and H4. Key regulatory steps involved in the determination of chromatin conformations are posttranslational modifications (PTM) at histone tails as well as the assembly of histone variants into nucleosomal arrays. Although the mechanistic background is fragmentary understood, it appears that the chromatin signature of metazoan cell types is inheritable over generations. Even less understood is the conservation of epigenetic mechanisms among eukaryotes and their origins.
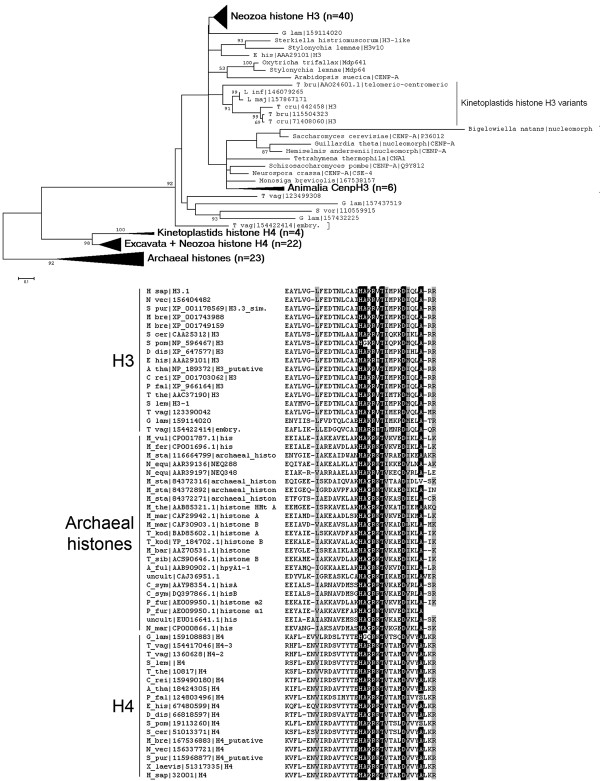
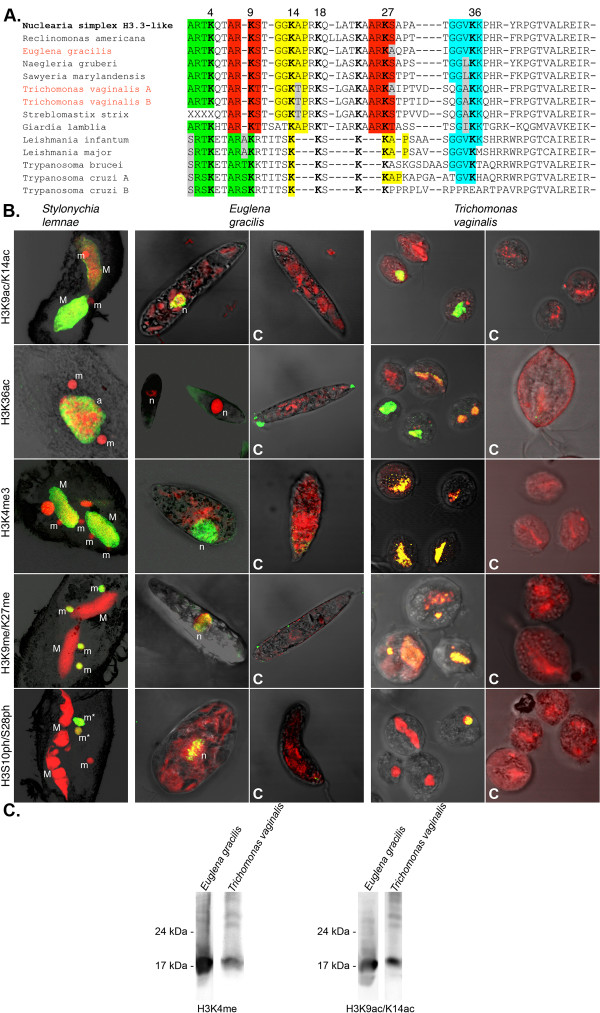
Results: In the light of recent progress in understanding the tree of eukaryotic life we discovered the origin of histone H3 by phylogenetic analyses of variants from all supergroups, which allowed the reconstruction of ancestral states. We found that H3 variants evolved frequently but independently within related species of almost all eukaryotic supergroups. Interestingly, we found all core histone types encoded in the genome of a basal dinoflagellate and H3 variants in two other species, although is was reported that dinoflagellate chromatin is not organized into nucleosomes.Most probably one or more animal/nuclearid H3.3-like variants gave rise to H3 variants of all opisthokonts (animals, choanozoa, fungi, nuclearids, Amoebozoa). H3.2 and H3.1 as well as H3.1t are derivatives of H3.3, whereas H3.2 evolved already in early branching animals, such as Trichoplax. H3.1 and H3.1t are probably restricted to mammals.We deduced a model for protoH3 of the last eukaryotic common ancestor (LECA) confirming a remarkable degree of sequence conservation in comparison to canonical human H3.1. We found evidence that multiple PTMs are conserved even in putatively early branching eukaryotic taxa (Euglenozoa/Excavata).
Conclusions: At least a basal repertoire of chromatin modifying mechanisms appears to share old common ancestry and may thus be inherent to all eukaryotes. We speculate that epigenetic principles responsive to environmental triggers may have had influenced phenotypic variation and concomitantly may potentially have had impact on eukaryotic diversification.
Figures



References
-
- Sandman K, Reeve JN. Chromosome packaging by archaeal histones. Adv Appl Microbiol. 2001;50:75–99. full_text. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
