

Vaccination of rhesus macaques with the anthrax vaccine adsorbed vaccine produces a serum antibody response that effectively neutralizes receptor-bound protective antigen in vitro
- PMID: 20739500
- PMCID: PMC2976102
- DOI: 10.1128/CVI.00174-10
Vaccination of rhesus macaques with the anthrax vaccine adsorbed vaccine produces a serum antibody response that effectively neutralizes receptor-bound protective antigen in vitro
Abstract
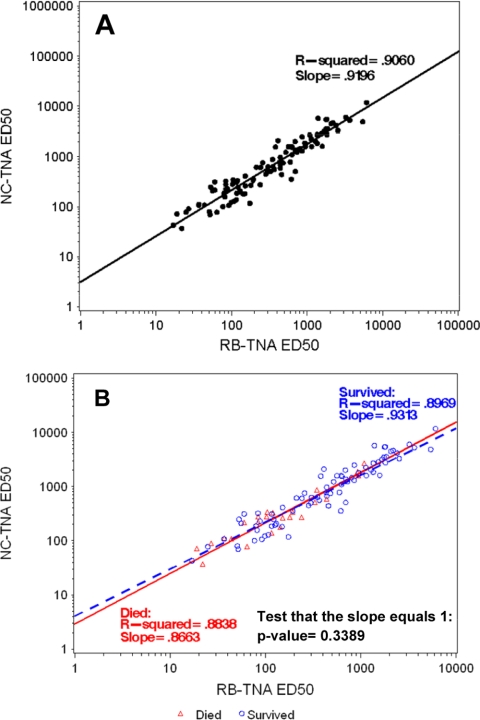
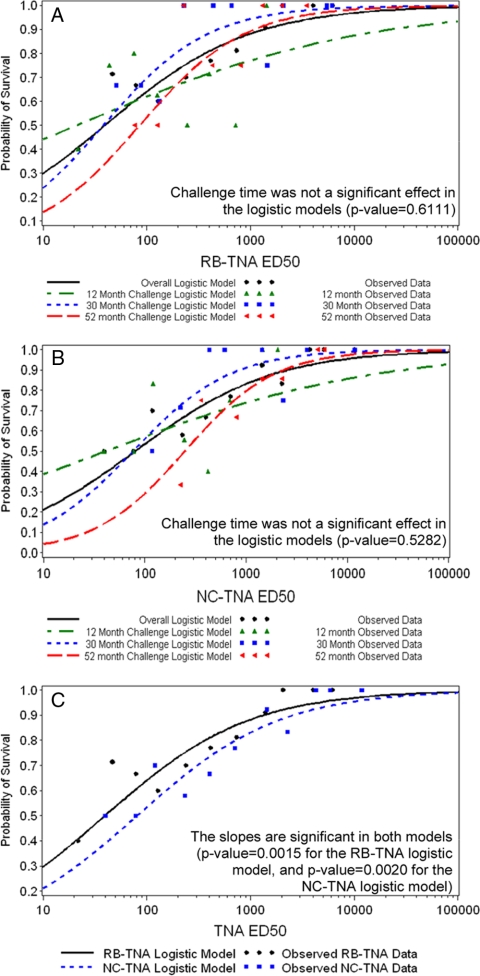
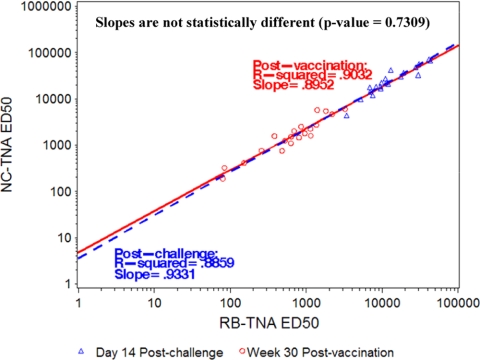
Anthrax toxin (ATx) is composed of the binary exotoxins lethal toxin (LTx) and edema toxin (ETx). They have separate effector proteins (edema factor and lethal factor) but have the same binding protein, protective antigen (PA). PA is the primary immunogen in the current licensed vaccine anthrax vaccine adsorbed (AVA [BioThrax]). AVA confers protective immunity by stimulating production of ATx-neutralizing antibodies, which could block the intoxication process at several steps (binding of PA to the target cell surface, furin cleavage, toxin complex formation, and binding/translocation of ATx into the cell). To evaluate ATx neutralization by anti-AVA antibodies, we developed two low-temperature LTx neutralization activity (TNA) assays that distinguish antibody blocking before and after binding of PA to target cells (noncomplexed [NC] and receptor-bound [RB] TNA assays). These assays were used to investigate anti-PA antibody responses in AVA-vaccinated rhesus macaques (Macaca mulatta) that survived an aerosol challenge with Bacillus anthracis Ames spores. Results showed that macaque anti-AVA sera neutralized LTx in vitro, even when PA was prebound to cells. Neutralization titers in surviving versus nonsurviving animals and between prechallenge and postchallenge activities were highly correlated. These data demonstrate that AVA stimulates a myriad of antibodies that recognize multiple neutralizing epitopes and confirm that change, loss, or occlusion of epitopes after PA is processed from PA83 to PA63 at the cell surface does not significantly affect in vitro neutralizing efficacy. Furthermore, these data support the idea that the full-length PA83 monomer is an appropriate immunogen for inclusion in next-generation anthrax vaccines.
Figures

Similar articles
-
A three-dose intramuscular injection schedule of anthrax vaccine adsorbed generates sustained humoral and cellular immune responses to protective antigen and provides long-term protection against inhalation anthrax in rhesus macaques.Clin Vaccine Immunol. 2012 Nov;19(11):1730-45. doi: 10.1128/CVI.00324-12. Epub 2012 Aug 29. Clin Vaccine Immunol. 2012. PMID: 22933399 Free PMC article.
-
Efficacy and immunogenicity of single-dose AdVAV intranasal anthrax vaccine compared to anthrax vaccine absorbed in an aerosolized spore rabbit challenge model.Clin Vaccine Immunol. 2015 Apr;22(4):430-9. doi: 10.1128/CVI.00690-14. Epub 2015 Feb 11. Clin Vaccine Immunol. 2015. PMID: 25673303 Free PMC article.
-
Comprehensive analysis and selection of anthrax vaccine adsorbed immune correlates of protection in rhesus macaques.Clin Vaccine Immunol. 2014 Nov;21(11):1512-20. doi: 10.1128/CVI.00469-14. Epub 2014 Sep 3. Clin Vaccine Immunol. 2014. PMID: 25185577 Free PMC article.
-
Neutralizing antibody and functional mapping of Bacillus anthracis protective antigen-The first step toward a rationally designed anthrax vaccine.Vaccine. 2016 Jan 2;34(1):13-9. doi: 10.1016/j.vaccine.2015.11.025. Epub 2015 Nov 21. Vaccine. 2016. PMID: 26611201 Review.
-
Molecular basis for improved anthrax vaccines.Adv Drug Deliv Rev. 2005 Jun 17;57(9):1266-92. doi: 10.1016/j.addr.2005.01.028. Epub 2005 Apr 21. Adv Drug Deliv Rev. 2005. PMID: 15935874 Review.
Cited by
-
Analysis of defined combinations of monoclonal antibodies in anthrax toxin neutralization assays and their synergistic action.Clin Vaccine Immunol. 2012 May;19(5):731-9. doi: 10.1128/CVI.05714-11. Epub 2012 Mar 21. Clin Vaccine Immunol. 2012. PMID: 22441391 Free PMC article.
-
MHC class II and non-MHC class II genes differentially influence humoral immunity to Bacillus anthracis lethal factor and protective antigen.Toxins (Basel). 2012 Dec;4(12):1451-67. doi: 10.3390/toxins4121451. Toxins (Basel). 2012. PMID: 23342680 Free PMC article.
-
Monoclonal antibody therapies against anthrax.Toxins (Basel). 2011 Aug;3(8):1004-19. doi: 10.3390/toxins3081004. Epub 2011 Aug 15. Toxins (Basel). 2011. PMID: 22069754 Free PMC article. Review.
-
Systematic Profiling of Full-Length Ig and TCR Repertoire Diversity in Rhesus Macaque through Long Read Transcriptome Sequencing.J Immunol. 2020 Jun 15;204(12):3434-3444. doi: 10.4049/jimmunol.1901256. Epub 2020 May 6. J Immunol. 2020. PMID: 32376650 Free PMC article.
-
Toxin-neutralizing antibodies elicited by naturally acquired cutaneous anthrax are elevated following severe disease and appear to target conformational epitopes.PLoS One. 2020 Apr 15;15(4):e0230782. doi: 10.1371/journal.pone.0230782. eCollection 2020. PLoS One. 2020. PMID: 32294093 Free PMC article.
References
-
- Auerbach, S., and G. G. Wright. 1955. Studies on immunity in anthrax. VI. Immunizing activity of protective antigen against various strains of Bacillus anthracis. J. Immunol. 75:129-133. - PubMed
-
- Belova, E. V., S. A. Dubilei, T. B. Kravchenko, A. V. Kolesnikov, M. I. Zakharova, and I. G. Shemiakin. 2004. Monoclonal antibodies to B. anthracis protective antigen are capable to neutralize and to enhance the anthrax lethal toxin action in vitro. Mol. Gen. Mikrobiol. Virusol. 2004:21-26. (In Russian.) - PubMed
-
- Belton, F. C., H. M. Darlow, and D. W. Henderson. 1956. The use of anthrax antigen to immunise man and monkey. Lancet 271:476-479. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
