

Defects in mitochondrial axonal transport and membrane potential without increased reactive oxygen species production in a Drosophila model of Friedreich ataxia
- PMID: 20739558
- PMCID: PMC2943153
- DOI: 10.1523/JNEUROSCI.0529-10.2010
Defects in mitochondrial axonal transport and membrane potential without increased reactive oxygen species production in a Drosophila model of Friedreich ataxia
Abstract
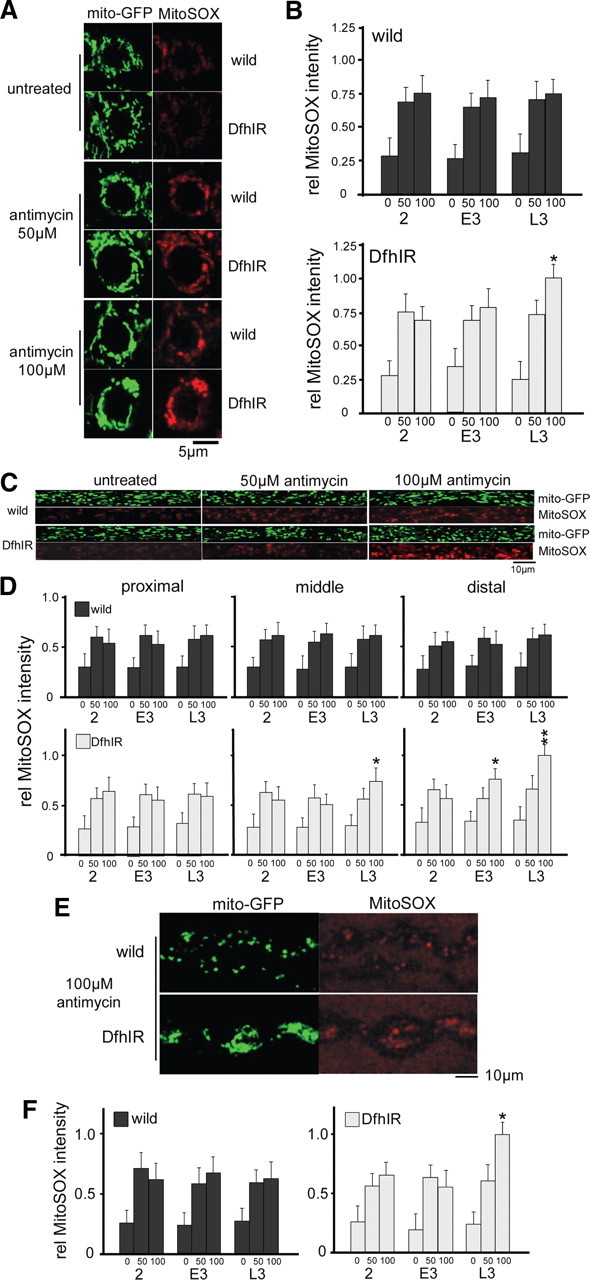
Friedreich ataxia, a neurodegenerative disorder resulting from frataxin deficiency, is thought to involve progressive cellular damage from oxidative stress. In Drosophila larvae with reduced frataxin expression (DfhIR), we evaluated possible mechanisms of cellular neuropathology by quantifying mitochondrial axonal transport, membrane potential (MMP), and reactive oxygen species (ROS) production in the DfhIR versus wild-type nervous system throughout development. Although dying-back neuropathy in DfhIR larvae did not occur until late third instar, reduced MMP was already apparent at second instar in the cell bodies, axons and neuromuscular junctions (NMJs) of segmental nerves. Defects in axonal transport of mitochondria appeared late in development in distal nerve of DfhIR larvae, with retrograde movement preferentially affected. As a result, by late third instar the neuromuscular junctions (NMJs) of DfhIR larvae accumulated a higher density of mitochondria, many of which were depolarized. Notably, increased ROS production was not detected in any neuronal region or developmental stage in DfhIR larvae. However, when challenged with antimycin A, neurons did respond with a larger increase in ROS. We propose that pathology in the frataxin-deficient nervous system involves decreased MMP and ATP production followed by failures of mitochondrial transport and NMJ function.
Figures

Similar articles
-
ROS regulation of axonal mitochondrial transport is mediated by Ca2+ and JNK in Drosophila.PLoS One. 2017 May 18;12(5):e0178105. doi: 10.1371/journal.pone.0178105. eCollection 2017. PLoS One. 2017. PMID: 28542430 Free PMC article.
-
Exenatide induces frataxin expression and improves mitochondrial function in Friedreich ataxia.JCI Insight. 2020 Jan 30;5(2):e134221. doi: 10.1172/jci.insight.134221. JCI Insight. 2020. PMID: 31877117 Free PMC article.
-
PPAR gamma agonist leriglitazone improves frataxin-loss impairments in cellular and animal models of Friedreich Ataxia.Neurobiol Dis. 2021 Jan;148:105162. doi: 10.1016/j.nbd.2020.105162. Epub 2020 Nov 7. Neurobiol Dis. 2021. PMID: 33171227
-
Oxidative stress and altered lipid metabolism in Friedreich ataxia.Free Radic Biol Med. 2016 Nov;100:138-146. doi: 10.1016/j.freeradbiomed.2016.06.007. Epub 2016 Jun 11. Free Radic Biol Med. 2016. PMID: 27296838 Review.
-
Iron metabolism and mitochondrial abnormalities in Friedreich ataxia.Blood Cells Mol Dis. 2002 Nov-Dec;29(3):536-47; discussion 548-52. doi: 10.1006/bcmd.2002.0591. Blood Cells Mol Dis. 2002. PMID: 12547248 Review.
Cited by
-
Overexpression of human and fly frataxins in Drosophila provokes deleterious effects at biochemical, physiological and developmental levels.PLoS One. 2011;6(7):e21017. doi: 10.1371/journal.pone.0021017. Epub 2011 Jul 11. PLoS One. 2011. PMID: 21779322 Free PMC article.
-
Charcot-Marie-Tooth disease and intracellular traffic.Prog Neurobiol. 2012 Dec;99(3):191-225. doi: 10.1016/j.pneurobio.2012.03.003. Epub 2012 Mar 22. Prog Neurobiol. 2012. PMID: 22465036 Free PMC article. Review.
-
The Organization of Mitochondrial Quality Control and Life Cycle in the Nervous System In Vivo in the Absence of PINK1.J Neurosci. 2015 Jun 24;35(25):9391-401. doi: 10.1523/JNEUROSCI.1198-15.2015. J Neurosci. 2015. PMID: 26109662 Free PMC article.
-
Drosophila melanogaster Models of Friedreich's Ataxia.Biomed Res Int. 2018 Apr 5;2018:5065190. doi: 10.1155/2018/5065190. eCollection 2018. Biomed Res Int. 2018. PMID: 29850527 Free PMC article. Review.
-
The role of oxidative stress in Friedreich's ataxia.FEBS Lett. 2018 Mar;592(5):718-727. doi: 10.1002/1873-3468.12928. Epub 2017 Dec 20. FEBS Lett. 2018. PMID: 29197070 Free PMC article. Review.
References
-
- Alexander C, Votruba M, Pesch UE, Thiselton DL, Mayer S, Moore A, Rodriguez M, Kellner U, Leo-Kottler B, Auburger G, Bhattacharya SS, Wissinger B. OPA1, encoding a dynamin-related GTPase, is mutated in autosomal dominant optic atrophy linked to chromosome 3q28. Nat Genet. 2000;26:211–215. - PubMed
-
- Anderson PR, Kirby K, Hilliker AJ, Phillips JP. RNAi-mediated suppression of the mitochondrial iron chaperone, frataxin, in Drosophila. Hum Mol Genet. 2005;14:3397–3405. - PubMed
-
- Balaban RS, Nemoto S, Finkel T. Mitochondria, oxidants, and aging. Cell. 2005;120:483–495. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials