

Auditory nerve fibers excite targets through synapses that vary in convergence, strength, and short-term plasticity
- PMID: 20739600
- PMCID: PMC3350034
- DOI: 10.1152/jn.00451.2010
Auditory nerve fibers excite targets through synapses that vary in convergence, strength, and short-term plasticity
Abstract
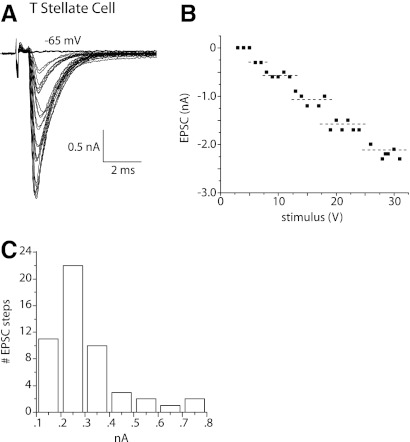
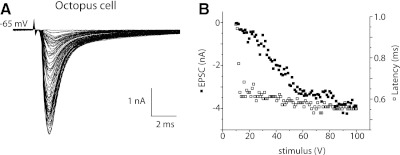
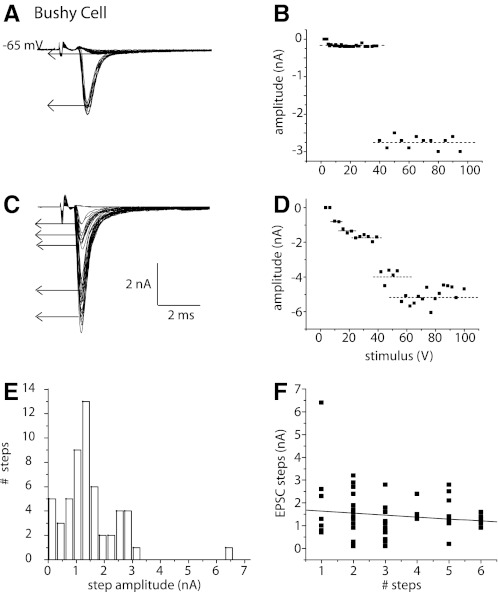
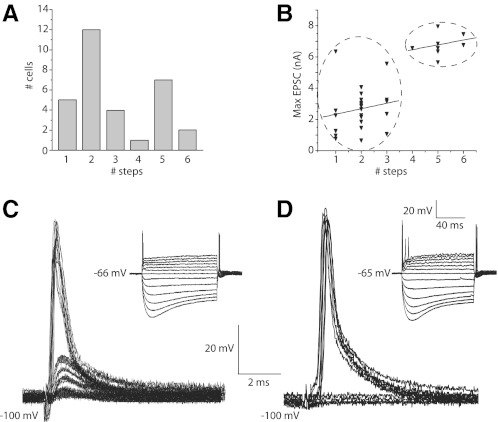
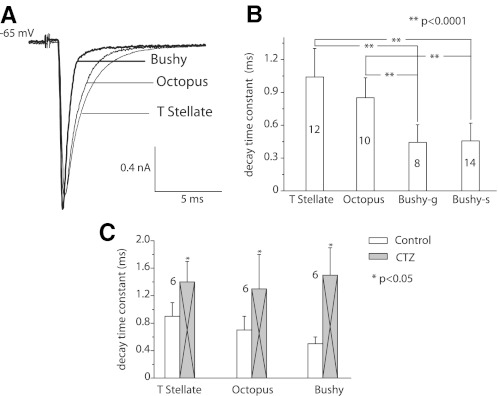
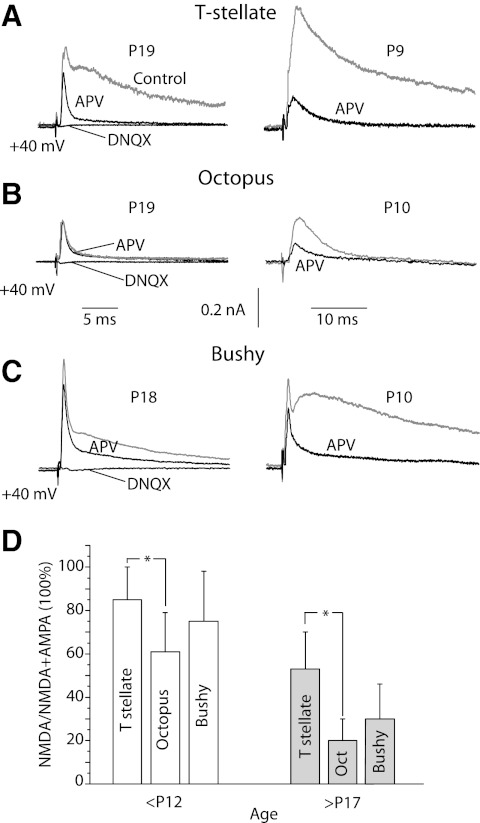
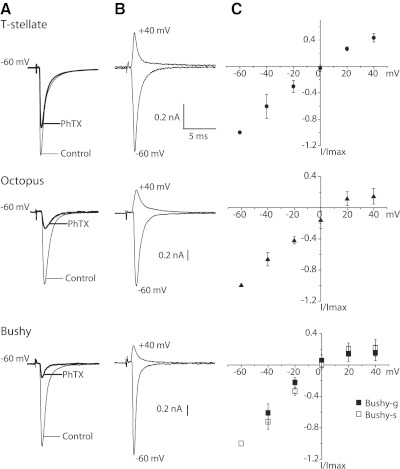
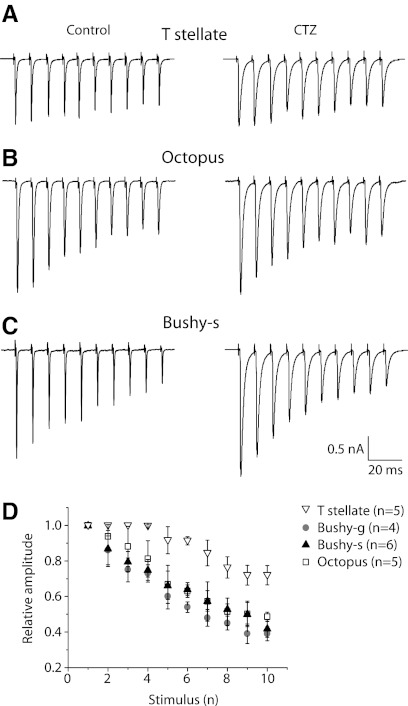
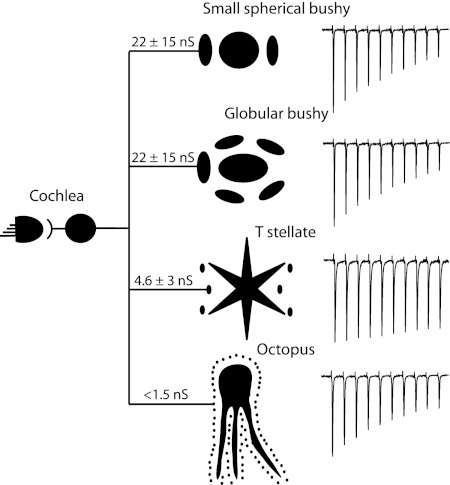
Auditory nerve fibers are the major source of excitation to the three groups of principal cells of the ventral cochlear nucleus (VCN), bushy, T stellate, and octopus cells. Shock-evoked excitatory postsynaptic currents (eEPSCs) in slices from mice showed systematic differences between groups of principal cells, indicating that target cells contribute to determining pre- and postsynaptic properties of synapses from spiral ganglion cells. Bushy cells likely to be small spherical bushy cells receive no more than three, most often two, excitatory inputs; those likely to be globular bushy cells receive at least four, most likely five, inputs. T stellate cells receive 6.5 inputs. Octopus cells receive >60 inputs. The N-methyl-d-aspartate (NMDA) components of eEPSCs were largest in T stellate, smaller in bushy, and smallest in octopus cells, and they were larger in neurons from younger than older mice. The average AMPA conductance of a unitary input is 22 ± 15 nS in both groups of bushy cells, <1.5 nS in octopus cells, and 4.6 ± 3 nS in T stellate cells. Sensitivity to philanthotoxin (PhTX) and rectification in the intracellular presence of spermine indicate that AMPA receptors that mediate eEPSCs in T stellate cells contain more GluR2 subunits than those in bushy and octopus cells. The AMPA components of eEPSCs were briefer in bushy (0.5 ms half-width) than in T stellate and octopus cells (0.8-0.9 ms half-width). Widening of eEPSCs in the presence of cyclothiazide (CTZ) indicates that desensitization shortens eEPSCs. CTZ-insensitive synaptic depression of the AMPA components was greater in bushy and octopus than in T stellate cells.
Figures
References
-
- Bal R, Baydas G, Naziroglu M. Electrophysiological properties of ventral cochlear nucleus neurons of the dog. Hear Res 256: 93–103, 2009 - PubMed
-
- Bal R, Oertel D. Hyperpolarization-activated, mixed-cation current (Ih) in octopus cells of the mammalian cochlear nucleus. J Neurophysiol 84: 806–817, 2000 - PubMed
-
- Bal R, Oertel D. Potassium currents in octopus cells of the mammalian cochlear nuclei. J Neurophysiol 86: 2299–2311, 2001 - PubMed
-
- Blackburn CC, Sachs MB. Classification of unit types in the anteroventral cochlear nucleus: PST histograms and regularity analysis. J Neurophysiol 62: 1303–1329, 1989 - PubMed
-
- Blackburn CC, Sachs MB. The representations of the steady-state vowel sound /e/ in the discharge patterns of cat anteroventral cochlear nucleus neurons. J Neurophysiol 63: 1191–1212, 1990 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous