

Functional elucidation of a key contact between tRNA and the large ribosomal subunit rRNA during decoding
- PMID: 20739608
- PMCID: PMC2941108
- DOI: 10.1261/rna.2232710
Functional elucidation of a key contact between tRNA and the large ribosomal subunit rRNA during decoding
Abstract
The selection of cognate tRNAs during translation is specified by a kinetic discrimination mechanism driven by distinct structural states of the ribosome. While the biochemical steps that drive the tRNA selection process have been carefully documented, it remains unclear how recognition of matched codon:anticodon helices in the small subunit facilitate global rearrangements in the ribosome complex that efficiently promote tRNA decoding. Here we use an in vitro selection approach to isolate tRNA(Trp) miscoding variants that exhibit a globally perturbed tRNA tertiary structure. Interestingly, the most substantial distortions are positioned in the elbow region of the tRNA that closely approaches helix 69 (H69) of the large ribosomal subunit. The importance of these specific interactions to tRNA selection is underscored by our kinetic analysis of both tRNA and rRNA variants that perturb the integrity of this interaction.
Figures
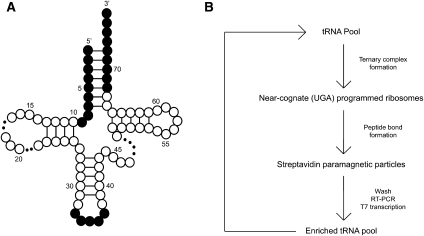
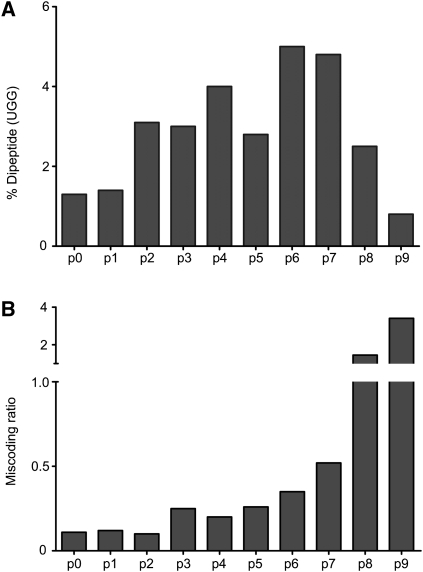
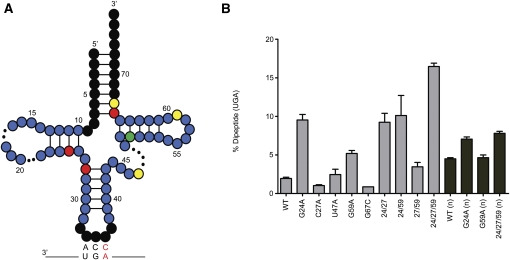
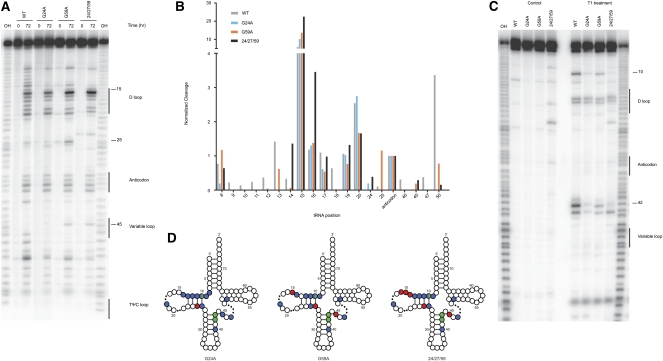
References
-
- Ali IK, Lancaster L, Feinberg J, Joseph S, Noller HF 2006. Deletion of a conserved, central ribosomal intersubunit RNA bridge. Mol Cell 23: 865–874 - PubMed
-
- Daviter T, Wieden HJ, Rodnina MV 2003. Essential role of histidine 84 in elongation factor Tu for the chemical step of GTP hydrolysis on the ribosome. J Mol Biol 332: 689–699 - PubMed
-
- Eisenberg SP, Soll L, Yarus M 1979. The purification and sequence of a temperature-sensitive tryptophan tRNA. J Biol Chem 254: 5562–5566 - PubMed
-
- Gromadski KB, Rodnina MV 2004. Kinetic determinants of high-fidelity tRNA discrimination on the ribosome. Mol Cell 13: 191–200 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources