

The poly(A)-binding protein partner Paip2a controls translation during late spermiogenesis in mice
- PMID: 20739757
- PMCID: PMC2929737
- DOI: 10.1172/JCI43350
The poly(A)-binding protein partner Paip2a controls translation during late spermiogenesis in mice
Abstract
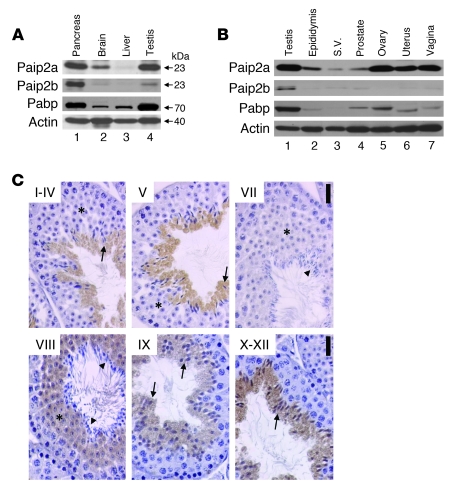
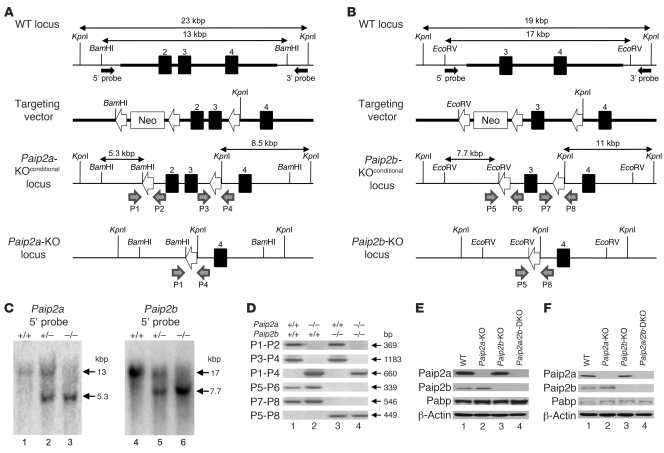
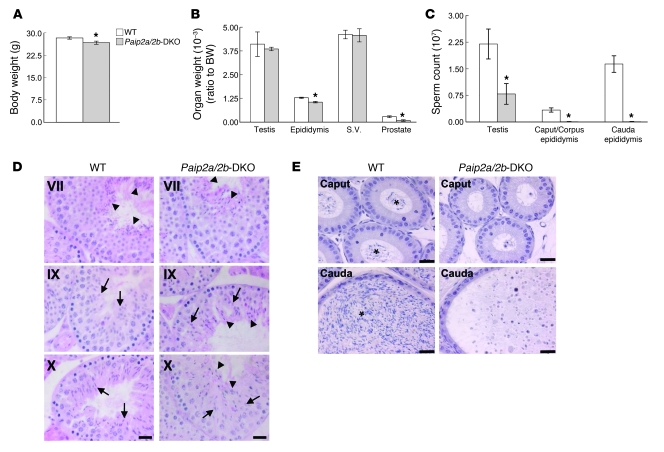
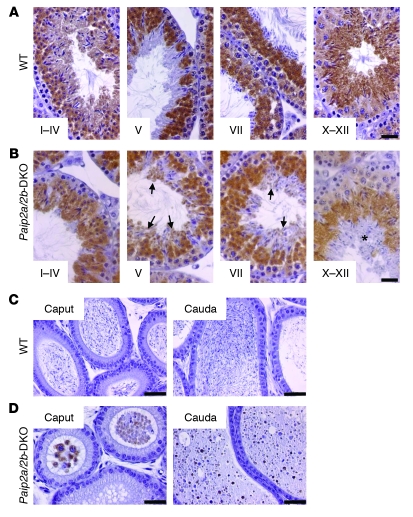
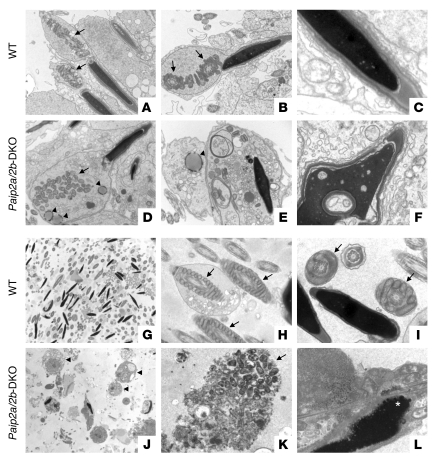
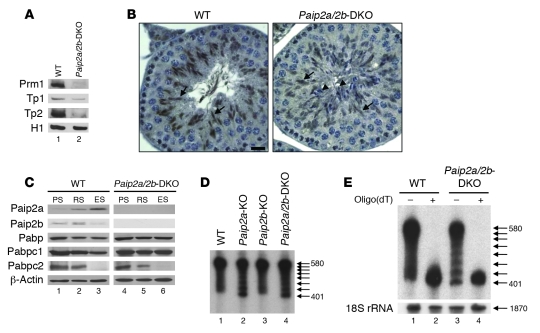
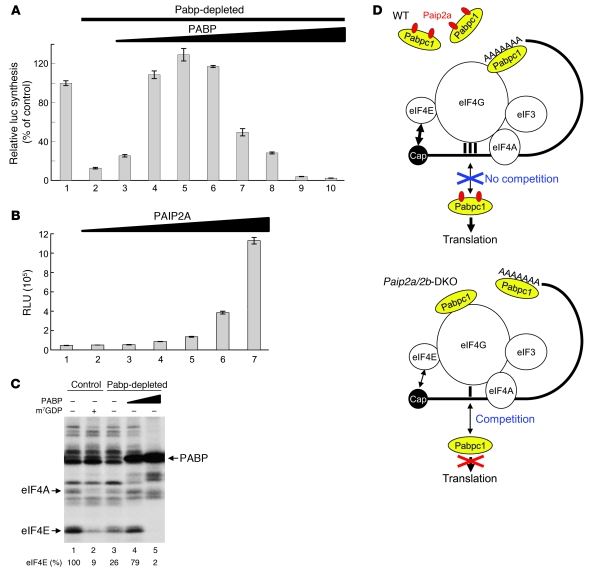
Translational control plays a key role in late spermiogenesis. A number of mRNAs encoding proteins required for late spermiogenesis are expressed in early spermatids but are stored as translationally inactive messenger ribonucleoprotein particles (mRNPs). The translation of these mRNAs is associated with shortening of their poly(A) tail in late spermiogenesis. Poly(A)-binding protein (Pabp) plays an important role in mRNA stabilization and translation. Three Pabp-interacting proteins, Paip1, Paip2a, and Paip2b, have been described. Paip2a is expressed in late spermatids. To investigate the role of Paip2 in spermiogenesis, we generated mice with knockout of either Paip2a or Paip2b and double-KO (DKO) mice lacking both Paip2a and Paip2b. Paip2a-KO and Paip2a/Paip2b-DKO mice exhibited male infertility. Translation of several mRNAs encoding proteins essential to male germ cell development was inhibited in late spermiogenesis in Paip2a/Paip2b-DKO mice, resulting in defective elongated spermatids. Inhibition of translation in Paip2a/Paip2b-DKO mice was caused by aberrant increased expression of Pabp, which impaired the interaction between eukaryotic initiation factor 4E (eIF4E) and the cap structure at the 5' end of the mRNA. We therefore propose a model whereby efficient mRNA translation in late spermiogenesis occurs at an optimal concentration of Pabp, a condition not fulfilled in Paip2a/Paip2b-DKO mice.
Figures
Comment in
-
Too much PABP, too little translation.J Clin Invest. 2010 Sep;120(9):3090-3. doi: 10.1172/JCI44091. Epub 2010 Aug 25. J Clin Invest. 2010. PMID: 20739750 Free PMC article.
References
-
- Kerr JB, Loveland KL, O’Bryan MK, de Kretser DM. Cytology of the testis and intrinsic control mechanisms. In: Neill JD, ed.Physiology of Reproduction . 3rd ed. Maryland Heights, Missouri, USA: Elsevier Press; 2006:1.
-
- Steger K. Transcriptional and translational regulation of gene expression in haploid spermatids. Anat Embryol (Berl). 1999;199(6):471–487. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
