

A model of yeast cell-cycle regulation based on multisite phosphorylation
- PMID: 20739927
- PMCID: PMC2947364
- DOI: 10.1038/msb.2010.55
A model of yeast cell-cycle regulation based on multisite phosphorylation
Abstract
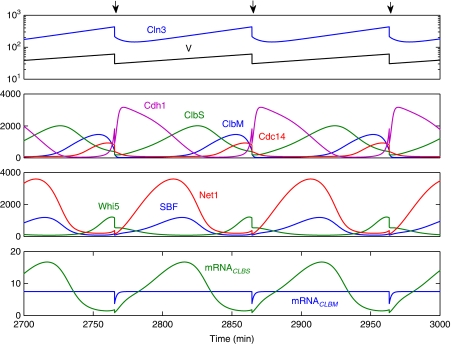
In order for the cell's genome to be passed intact from one generation to the next, the events of the cell cycle (DNA replication, mitosis, cell division) must be executed in the correct order, despite the considerable molecular noise inherent in any protein-based regulatory system residing in the small confines of a eukaryotic cell. To assess the effects of molecular fluctuations on cell-cycle progression in budding yeast cells, we have constructed a new model of the regulation of Cln- and Clb-dependent kinases, based on multisite phosphorylation of their target proteins and on positive and negative feedback loops involving the kinases themselves. To account for the significant role of noise in the transcription and translation steps of gene expression, the model includes mRNAs as well as proteins. The model equations are simulated deterministically and stochastically to reveal the bistable switching behavior on which proper cell-cycle progression depends and to show that this behavior is robust to the level of molecular noise expected in yeast-sized cells (approximately 50 fL volume). The model gives a quantitatively accurate account of the variability observed in the G1-S transition in budding yeast, which is governed by an underlying sizer+timer control system.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures


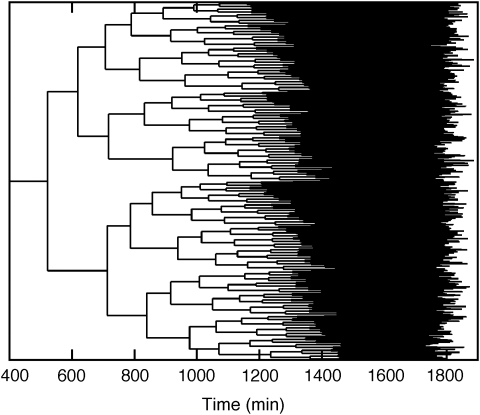






 ploidy. Left panel: daughter cells; right panel: mother cells, except for last row.
ploidy. Left panel: daughter cells; right panel: mother cells, except for last row.

References
-
- Amon A, Tyers M, Futcher B, Nasmyth K (1993) Mechanisms that help the yeast cell cycle clock tick: G2 cyclins transcriptionally activate G2 cyclins and repress G1 cyclins. Cell 74: 993–1007 - PubMed
-
- Azzam R, Chen SL, Shou WY, Mah AS, Alexandru G, Nasmyth K, Annan RS, Carr SA, Deshaies RJ (2004) Phosphorylation by cyclin B-CDK underlies release of mitotic exit activator Cdc14 from the nucleolus. Science 305: 516–519 - PubMed
-
- Braunewell S, Bornholdt S (2007) Superstability of the yeast cell-cycle dynamics: ensuring causality in the presence of biochemical stochasticity. J Theor Biol 245: 638–643 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
