

Probing the natural scene by echolocation in bats
- PMID: 20740076
- PMCID: PMC2927269
- DOI: 10.3389/fnbeh.2010.00033
Probing the natural scene by echolocation in bats
Abstract
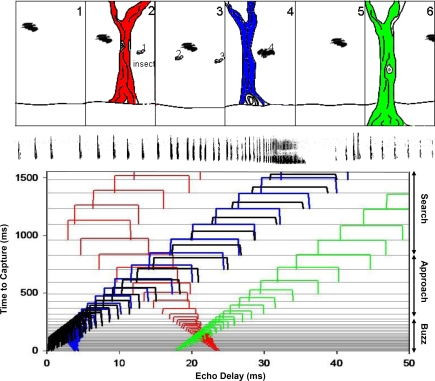
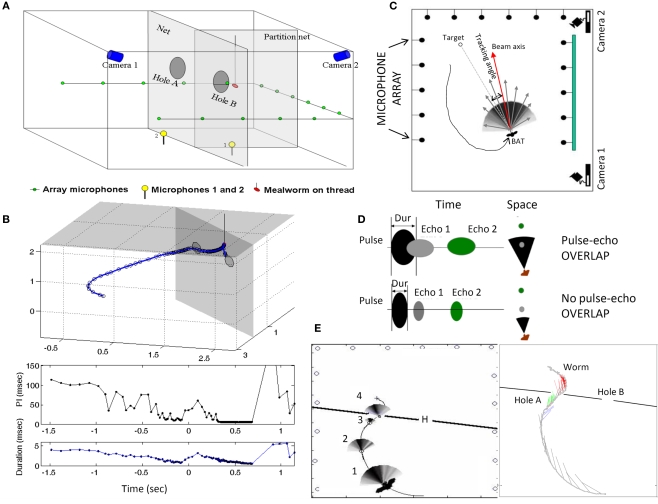
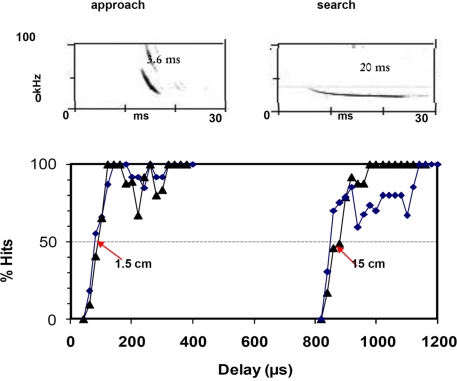
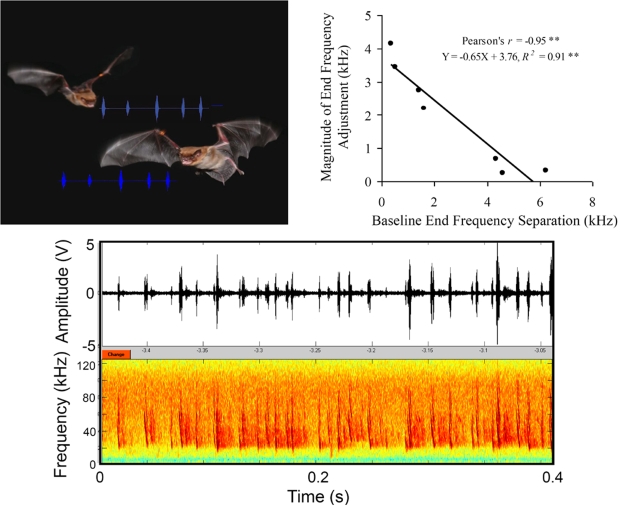
Bats echolocating in the natural environment face the formidable task of sorting signals from multiple auditory objects, echoes from obstacles, prey, and the calls of conspecifics. Successful orientation in a complex environment depends on auditory information processing, along with adaptive vocal-motor behaviors and flight path control, which draw upon 3-D spatial perception, attention, and memory. This article reviews field and laboratory studies that document adaptive sonar behaviors of echolocating bats, and point to the fundamental signal parameters they use to track and sort auditory objects in a dynamic environment. We suggest that adaptive sonar behavior provides a window to bats' perception of complex auditory scenes.
Keywords: action-perception; auditory scene analysis; hearing; neuroethology; stream segregation.
Figures

References
-
- Au W. W. L. (1993). The Sonar of Dolphins. New York: Springer
-
- Aytekin M., Grassi E., Sahota M., Moss C. F. (2004). The bat head-related transfer function reveals binaural cues for sound. - PubMed
-
- Barak Y., Yom-Tov Y. (1989). The advantage of group hunting to Kuhl's pipistrelle bat Pipistrellus kuhli (Microchiroptera). J. Zool. 219, 670–675 10.1111/j.1469-7998.1989.tb02608.x - DOI
LinkOut - more resources
Full Text Sources