

Substitution of Ala for Tyr567 in RB69 DNA polymerase allows dAMP and dGMP to be inserted opposite Guanidinohydantoin
- PMID: 20795733
- PMCID: PMC3755731
- DOI: 10.1021/bi100913v
Substitution of Ala for Tyr567 in RB69 DNA polymerase allows dAMP and dGMP to be inserted opposite Guanidinohydantoin
Abstract
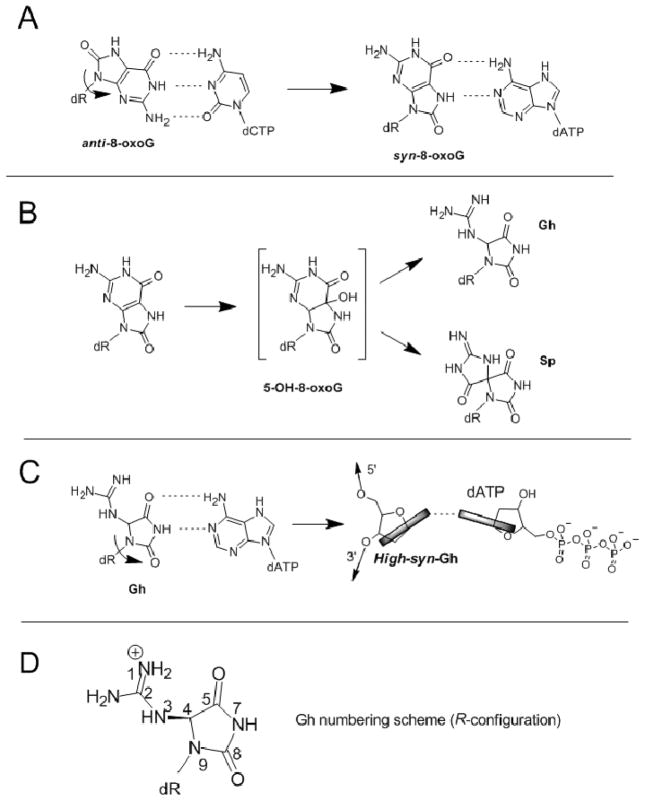
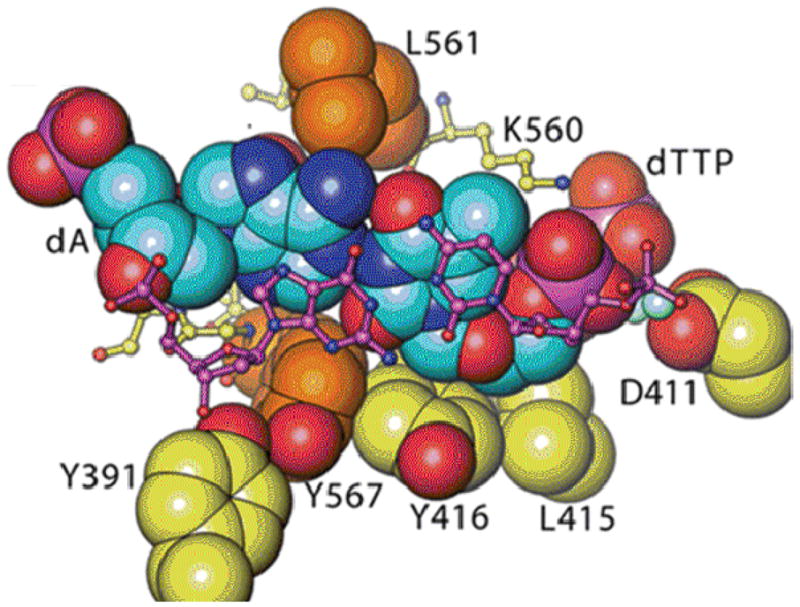
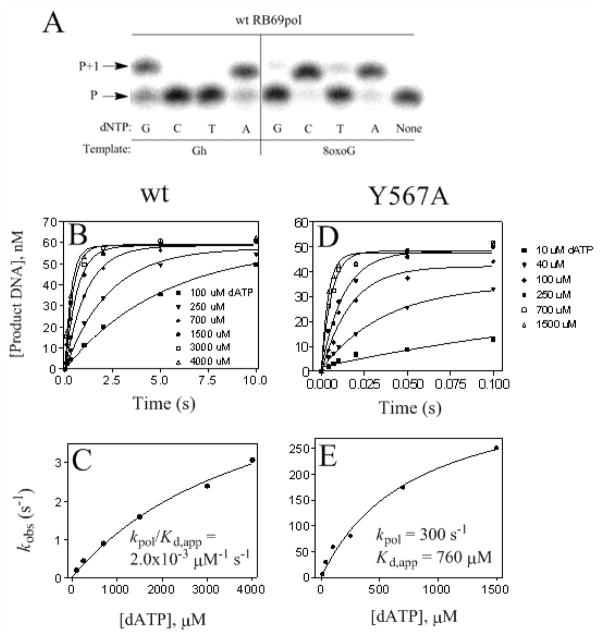
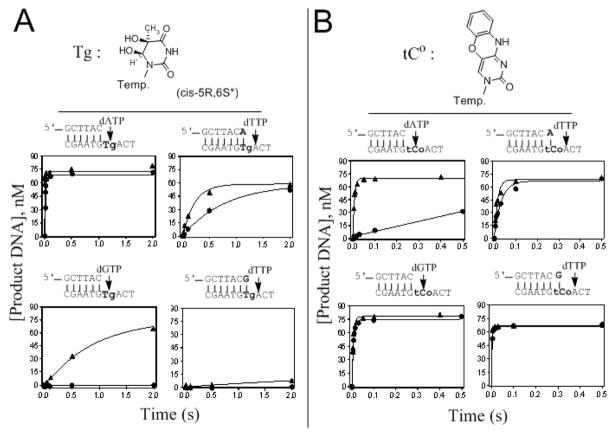
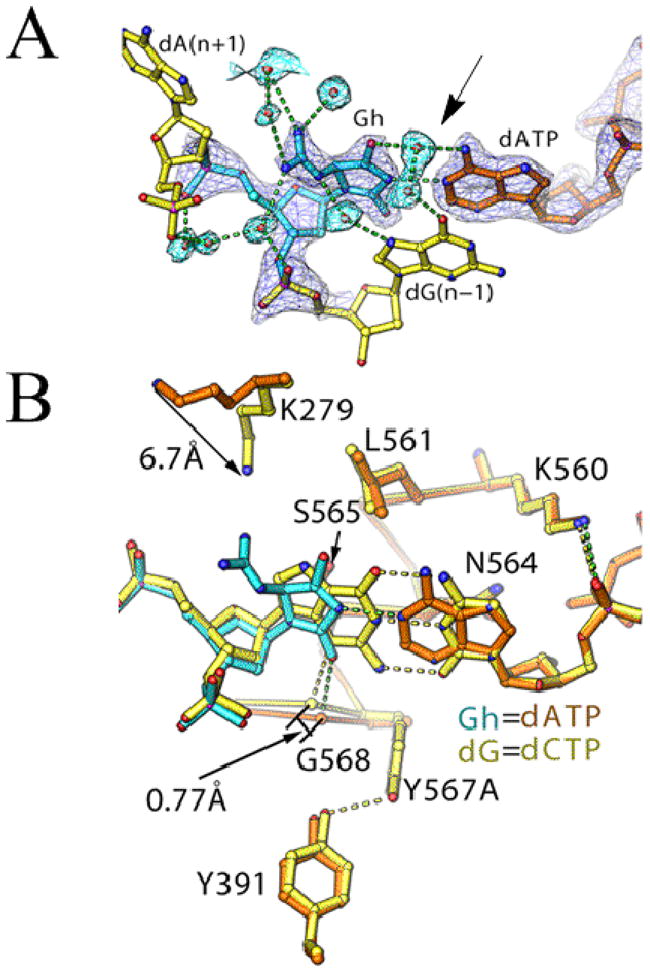
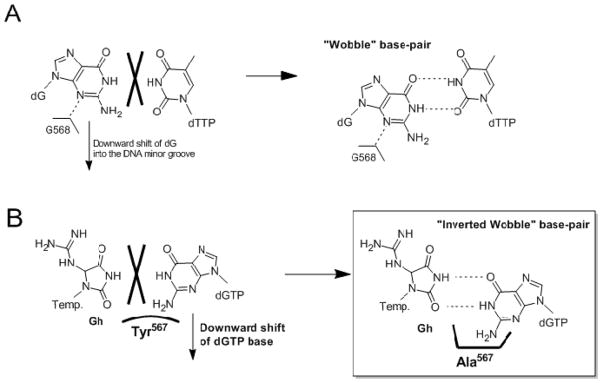
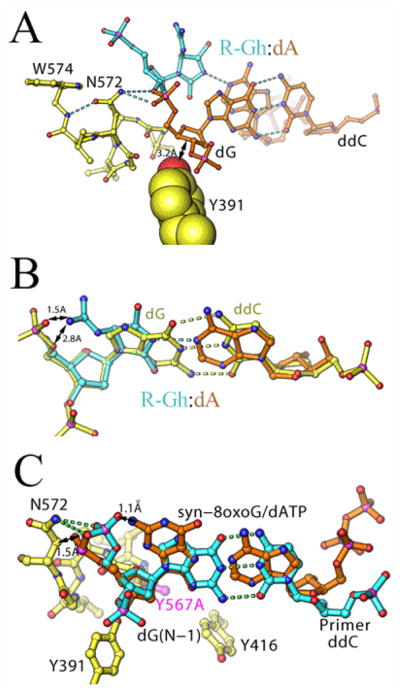
Continuous oxidative damage inflicted on DNA produces 7,8-dihydro-8-oxoguanine (8-oxoG), a commonly occurring lesion that can potentially cause cancer by producing G → T transversions during DNA replication. Mild oxidation of 8-oxoG leads to the formation of hydantoins, specifically guanidinohydantoin (Gh) and spiroiminodihydantoin (Sp), which are 100% mutagenic because they encode almost exclusively the insertion of dAMP and dGMP (encoding G → T and G → C transversions, respectively). The wild-type (wt) pol α family DNA polymerase from bacteriophage RB69 (RB69pol) inserts dAMP and dGMP with low efficiency when situated opposite Gh. In contrast, the RB69pol Y567A mutant inserts both of these dNMPs opposite Gh with >100-fold higher efficiency than wt. We now report the crystal structure of the "closed" preinsertion complex for the Y567A mutant with dATP opposite a templating Gh (R-configuration) in a 13/18mer primer-template (P/T) at 2.0 Å resolution. The structure data reveal that the Y to A substitution provides the nascent base pair binding pocket (NBP) with the flexibility to accommodate Gh by allowing G568 to move in the major-to-minor groove direction of the P/T. Thus, Gh is rejected as a templating base by wt RB69pol because G568 is inflexible, preventing Gh from pairing with the incoming dATP or dGTP base.
Figures
Similar articles
-
RB69 DNA polymerase structure, kinetics, and fidelity.Biochemistry. 2014 May 6;53(17):2752-67. doi: 10.1021/bi4014215. Epub 2014 Apr 23. Biochemistry. 2014. PMID: 24720884 Free PMC article. Review.
-
Using a fluorescent cytosine analogue tC(o) to probe the effect of the Y567 to Ala substitution on the preinsertion steps of dNMP incorporation by RB69 DNA polymerase.Biochemistry. 2012 Jun 5;51(22):4609-17. doi: 10.1021/bi300241m. Epub 2012 May 22. Biochemistry. 2012. PMID: 22616982 Free PMC article.
-
Substitution of Ala for Tyr567 in RB69 DNA polymerase allows dAMP to be inserted opposite 7,8-dihydro-8-oxoguanine.Biochemistry. 2010 May 18;49(19):4116-25. doi: 10.1021/bi100102s. Biochemistry. 2010. PMID: 20411947 Free PMC article.
-
Crystal structure of a replicative DNA polymerase bound to the oxidized guanine lesion guanidinohydantoin.Biochemistry. 2010 Mar 23;49(11):2502-9. doi: 10.1021/bi902195p. Biochemistry. 2010. PMID: 20166752 Free PMC article.
-
Utility of the bacteriophage RB69 polymerase gp43 as a surrogate enzyme for herpesvirus orthologs.Viruses. 2013 Jan 8;5(1):54-86. doi: 10.3390/v5010054. Viruses. 2013. PMID: 23299784 Free PMC article. Review.
Cited by
-
RB69 DNA polymerase structure, kinetics, and fidelity.Biochemistry. 2014 May 6;53(17):2752-67. doi: 10.1021/bi4014215. Epub 2014 Apr 23. Biochemistry. 2014. PMID: 24720884 Free PMC article. Review.
-
pH-Dependent Equilibrium between 5-Guanidinohydantoin and Iminoallantoin Affects Nucleotide Insertion Opposite the DNA Lesion.J Org Chem. 2016 Jan 15;81(2):351-9. doi: 10.1021/acs.joc.5b02180. Epub 2015 Nov 24. J Org Chem. 2016. PMID: 26582419 Free PMC article.
-
Calculation of the stabilization energies of oxidatively damaged guanine base pairs with guanine.Molecules. 2012 Jun 1;17(6):6705-15. doi: 10.3390/molecules17066705. Molecules. 2012. PMID: 22728364 Free PMC article.
-
Using a fluorescent cytosine analogue tC(o) to probe the effect of the Y567 to Ala substitution on the preinsertion steps of dNMP incorporation by RB69 DNA polymerase.Biochemistry. 2012 Jun 5;51(22):4609-17. doi: 10.1021/bi300241m. Epub 2012 May 22. Biochemistry. 2012. PMID: 22616982 Free PMC article.
-
Products of Oxidative Guanine Damage Form Base Pairs with Guanine.Int J Mol Sci. 2020 Oct 15;21(20):7645. doi: 10.3390/ijms21207645. Int J Mol Sci. 2020. PMID: 33076559 Free PMC article. Review.
References
-
- Steenken S, Jovanovic SV. How Easily Oxidizable Is DNA?. One-Electron Reduction Potentials of Adenosine and Guanosine Radicals in Aqueous Solution. J Am Chem Soc. 1997;119:617–618.
-
- Sekiguchi M, Tsuzuki T. Oxidative nucleotide damage: consequences and prevention. Oncogene. 2002;21:8895–8904. - PubMed
-
- Malins DC, Haimanot R. Major alterations in the nucleotide structure of DNA in cancer of the female breast. Cancer Res. 1991;51:5430–5432. - PubMed
-
- Degan P, Shigenaga MK, Park EM, Alperin PE, Ames BN. Immunoaffinity isolation of urinary 8-hydroxy-2′-deoxyguanosine and 8-hydroxyguanine and quantitation of 8-hydroxy-2′-deoxyguanosine in DNA by polyclonal antibodies. Carcinogenesis. 1991;12:865–871. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
