

Biliary innate immunity: function and modulation
- PMID: 20798866
- PMCID: PMC2926654
- DOI: 10.1155/2010/373878
Biliary innate immunity: function and modulation
Abstract
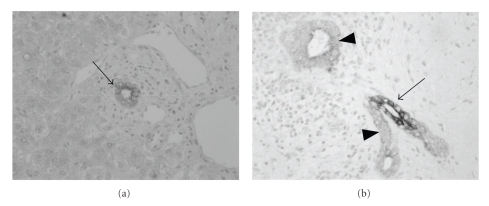
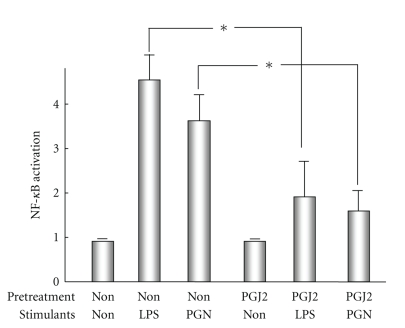
Biliary innate immunity is involved in the pathogenesis of cholangiopathies in patients with primary biliary cirrhosis (PBC) and biliary atresia. Biliary epithelial cells possess an innate immune system consisting of the Toll-like receptor (TLR) family and recognize pathogen-associated molecular patterns (PAMPs). Tolerance to bacterial PAMPs such as lipopolysaccharides is also important to maintain homeostasis in the biliary tree, but tolerance to double-stranded RNA (dsRNA) is not found. In PBC, CD4-positive Th17 cells characterized by the secretion of IL-17 are implicated in the chronic inflammation of bile ducts and the presence of Th17 cells around bile ducts is causally associated with the biliary innate immune responses to PAMPs. Moreover, a negative regulator of intracellular TLR signaling, peroxisome proliferator-activated receptor-gamma (PPARgamma), is involved in the pathogenesis of cholangitis. Immunosuppression using PPARgamma ligands may help to attenuate the bile duct damage in PBC patients. In biliary atresia characterized by a progressive, inflammatory, and sclerosing cholangiopathy, dsRNA viruses are speculated to be an etiological agent and to directly induce enhanced biliary apoptosis via the expression of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). Moreover, the epithelial-mesenchymal transition (EMT) of biliary epithelial cells is also evoked by the biliary innate immune response to dsRNA.
Figures
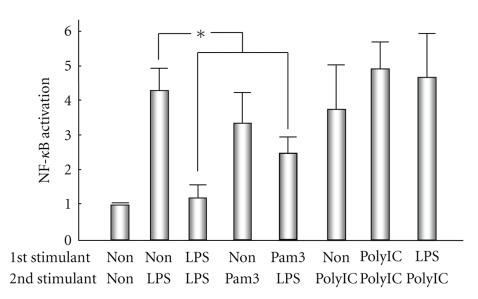
Similar articles
-
Biliary innate immunity in the pathogenesis of biliary diseases.Inflamm Allergy Drug Targets. 2010 Jun;9(2):83-90. doi: 10.2174/187152810791292809. Inflamm Allergy Drug Targets. 2010. PMID: 20402650 Review.
-
Th1 cytokine-induced downregulation of PPARgamma in human biliary cells relates to cholangitis in primary biliary cirrhosis.Hepatology. 2005 Jun;41(6):1329-38. doi: 10.1002/hep.20705. Hepatology. 2005. PMID: 15880426
-
Innate immunity in the pathogenesis of cholangiopathy: a recent update.Inflamm Allergy Drug Targets. 2012 Dec;11(6):478-83. doi: 10.2174/187152812803589976. Inflamm Allergy Drug Targets. 2012. PMID: 22920631 Review.
-
Periductal interleukin-17 production in association with biliary innate immunity contributes to the pathogenesis of cholangiopathy in primary biliary cirrhosis.Clin Exp Immunol. 2009 Aug;157(2):261-70. doi: 10.1111/j.1365-2249.2009.03947.x. Clin Exp Immunol. 2009. PMID: 19604266 Free PMC article.
-
Sclerosing and obstructive cholangiopathy in biliary atresia: mechanisms and association with biliary innate immunity.Pediatr Surg Int. 2017 Dec;33(12):1243-1248. doi: 10.1007/s00383-017-4154-8. Epub 2017 Oct 16. Pediatr Surg Int. 2017. PMID: 29039048 Review.
Cited by
-
Toll-like receptors in secondary obstructive cholangiopathy.Gastroenterol Res Pract. 2011;2011:265093. doi: 10.1155/2011/265093. Epub 2011 Oct 27. Gastroenterol Res Pract. 2011. PMID: 22114589 Free PMC article.
-
Liver fibrosis in biliary atresia.World J Pediatr. 2019 Apr;15(2):117-123. doi: 10.1007/s12519-018-0203-1. Epub 2018 Nov 21. World J Pediatr. 2019. PMID: 30465125 Review.
-
The Role of Microbiota in Liver Transplantation and Liver Transplantation-Related Biliary Complications.Int J Mol Sci. 2023 Mar 2;24(5):4841. doi: 10.3390/ijms24054841. Int J Mol Sci. 2023. PMID: 36902269 Free PMC article. Review.
-
Methotrexate-loaded tumour-cell-derived microvesicles can relieve biliary obstruction in patients with extrahepatic cholangiocarcinoma.Nat Biomed Eng. 2020 Jul;4(7):743-753. doi: 10.1038/s41551-020-0583-0. Epub 2020 Jul 6. Nat Biomed Eng. 2020. PMID: 32632227
-
Immune-mediated cholangiopathies in children: the need to better understand the pathophysiology for finding the future possible treatment targets.Front Immunol. 2023 Oct 20;14:1206025. doi: 10.3389/fimmu.2023.1206025. eCollection 2023. Front Immunol. 2023. PMID: 37928553 Free PMC article. Review.
References
-
- Lemaitre B, Nicolas E, Michaut L, Reichhart J-M, Hoffmann JA. The dorsoventral regulatory gene cassette spatzle/Toll/Cactus controls the potent antifungal response in Drosophila adults. Cell. 1996;86(6):973–983. - PubMed
-
- Akira S, Takeda K. Toll-like receptor signalling. Nature Reviews Immunology. 2004;4(7):499–511. - PubMed
-
- Hugot J-P, Chamaillard M, Zouali H, et al. Association of NOD2 leucine-rich repeat variants with susceptibility to Crohn’s disease. Nature. 2001;411(6837):599–603. - PubMed
-
- Harada K, Ohira S, Isse K, et al. Lipopolysaccharide activates nuclear factor-kappaB through Toll-like receptors and related molecules in cultured biliary epithelial cells. Laboratory Investigation. 2003;83(11):1657–1667. - PubMed
-
- Hiramatsu K, Harada K, Tsuneyama K, et al. Amplification and sequence analysis of partial bacterial 16S ribosomal RNA gene in gallbladder bile from patients with primary biliary cirrhosis. Journal of Hepatology. 2000;33(1):9–18. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
