

Ferricytochrome c protects mitochondrial cytochrome c oxidase against hydrogen peroxide-induced oxidative damage
- PMID: 20801213
- PMCID: PMC2953960
- DOI: 10.1016/j.freeradbiomed.2010.08.019
Ferricytochrome c protects mitochondrial cytochrome c oxidase against hydrogen peroxide-induced oxidative damage
Abstract
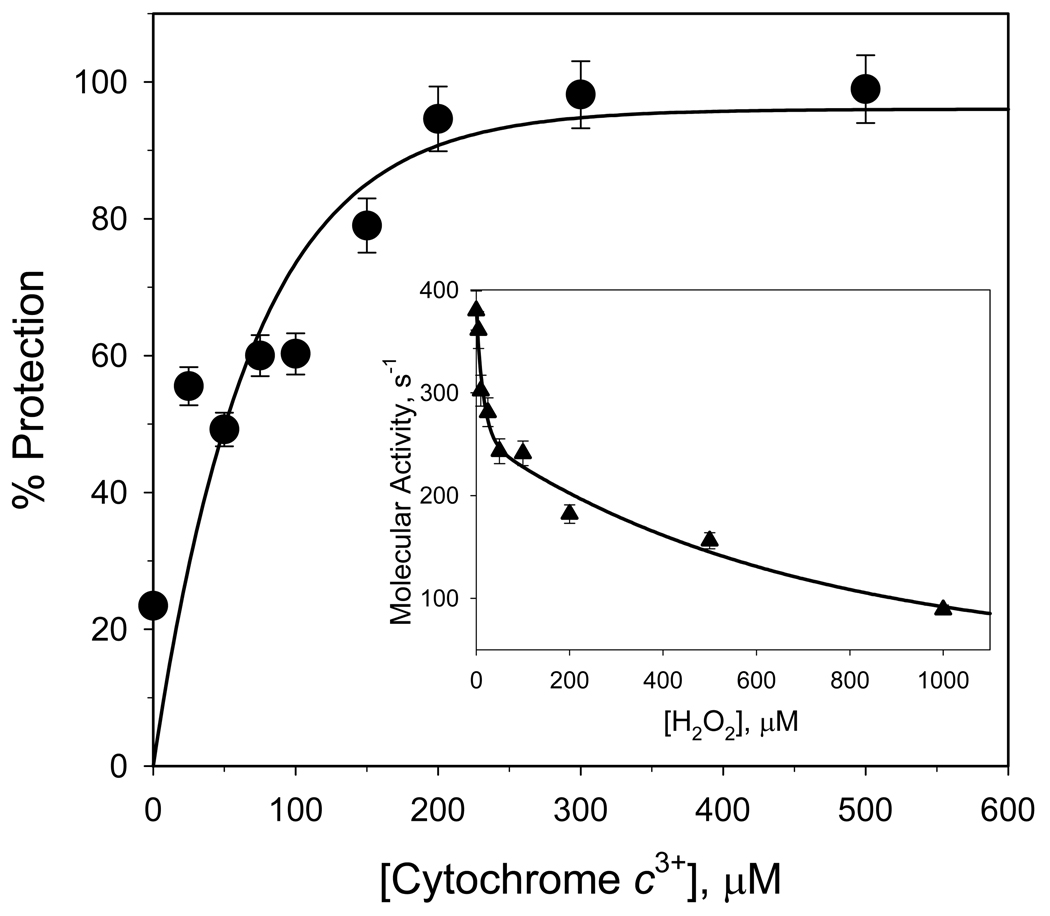
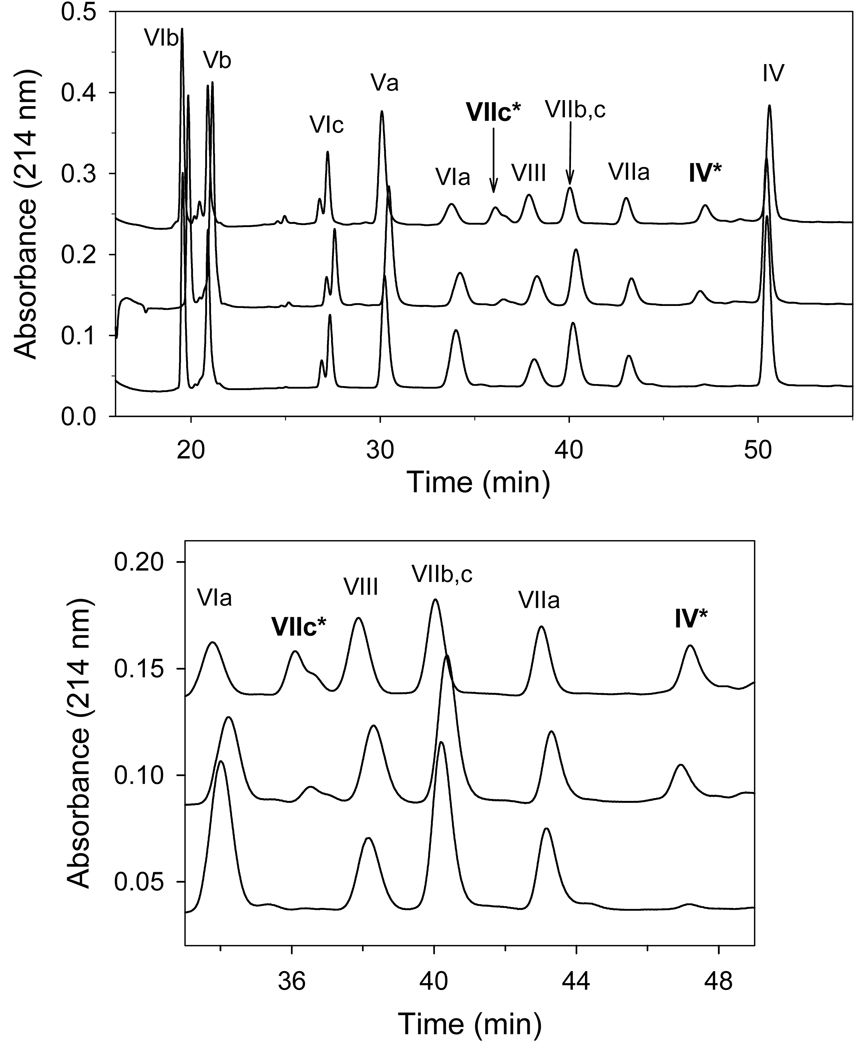
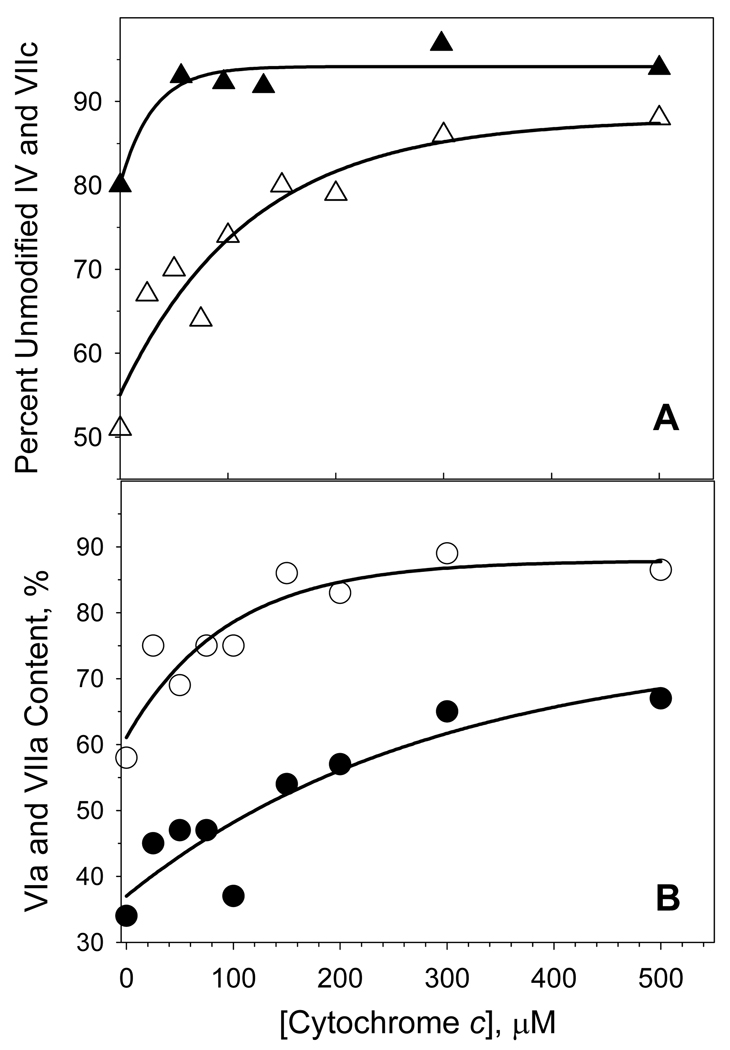
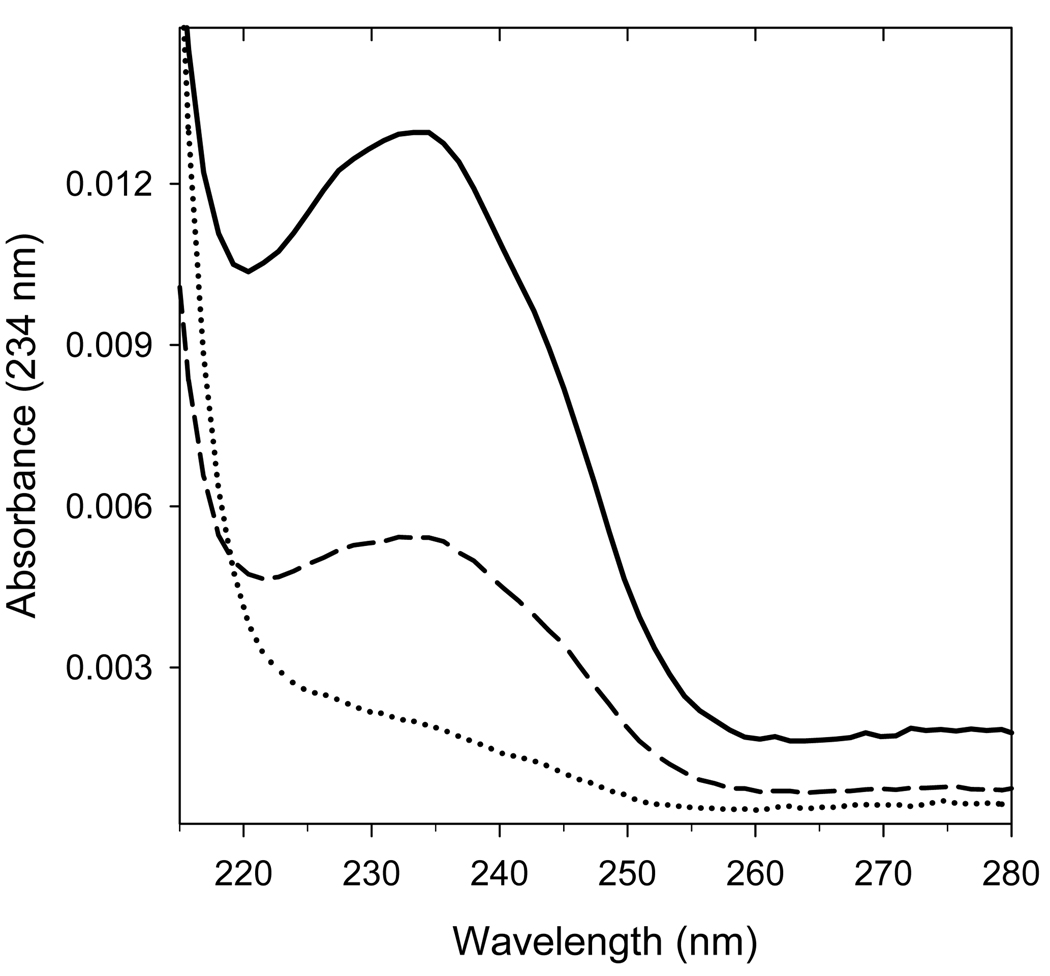
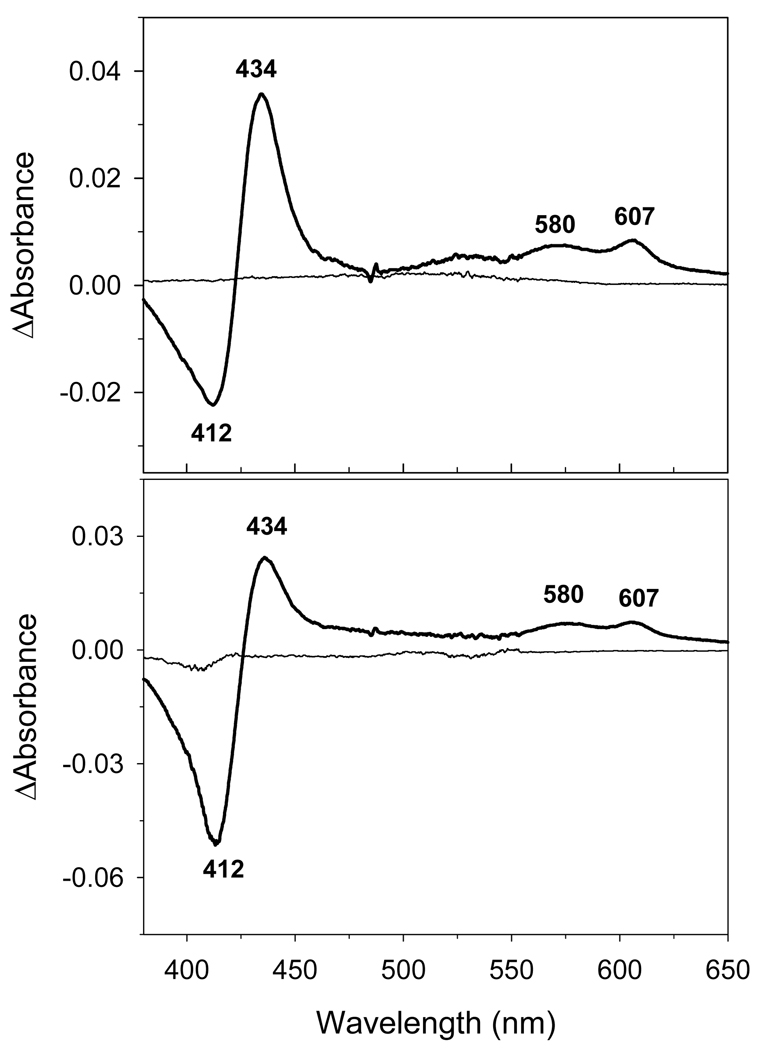
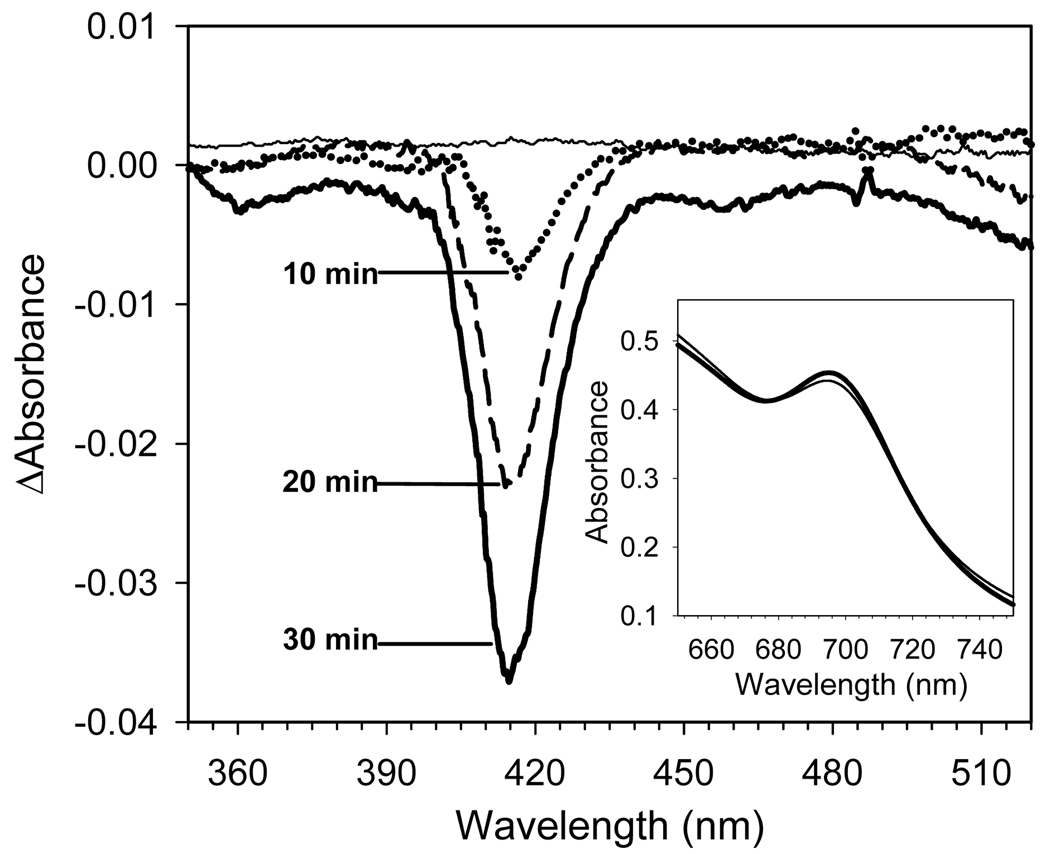
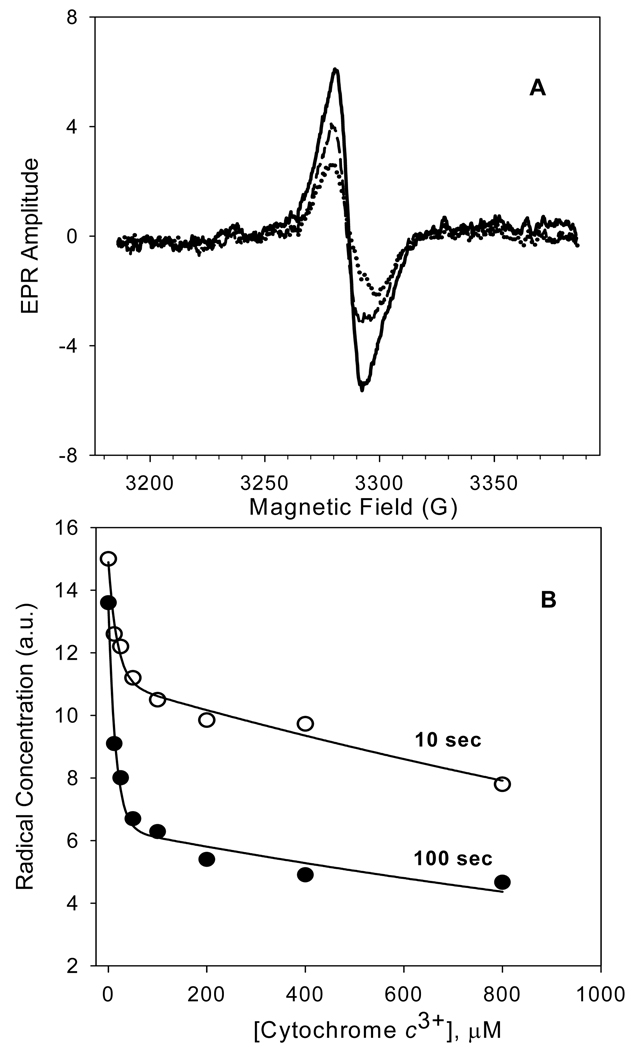
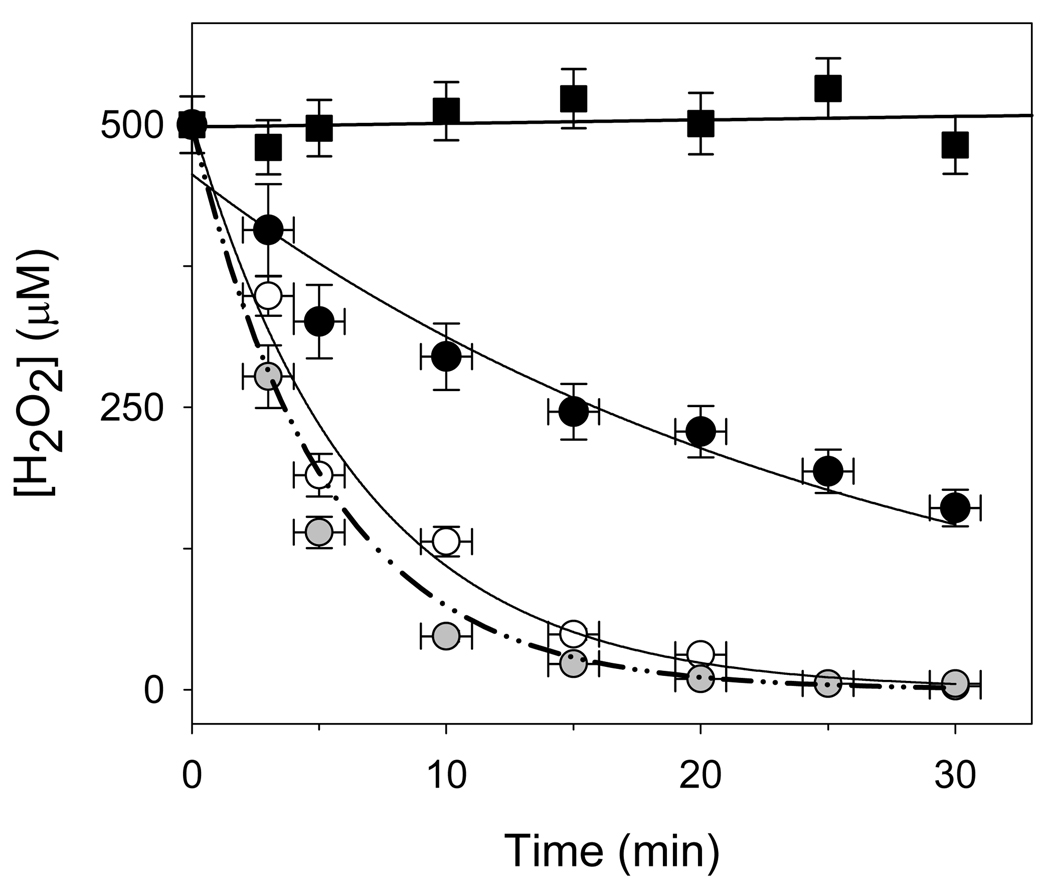
An excess of ferricytochrome c protects purified mitochondrial cytochrome c oxidase and bound cardiolipin from hydrogen peroxide-induced oxidative modification. All of the peroxide-induced changes within cytochrome c oxidase, such as oxidation of Trp(19,IV) and Trp(48,VIIc), partial dissociation of subunits VIa and VIIa, and generation of cardiolipin hydroperoxide, no longer take place in the presence of ferricytochrome c. Furthermore, ferricytochrome c suppresses the yield of H(2)O(2)-induced free radical detectable by electron paramagnetic resonance spectroscopy within cytochrome c oxidase. These protective effects are based on two mechanisms. The first involves the peroxidase/catalase-like activity of ferricytochrome c, which results in the decomposition of H(2)O(2), with the apparent bimolecular rate constant of 5.1±1.0M(-1)s(-1). Although this value is lower than the rate constant of a specialized peroxidase, the activity is sufficient to eliminate H(2)O(2)-induced damage to cytochrome c oxidase in the presence of an excess of ferricytochrome c. The second mechanism involves ferricytochrome c-induced quenching of free radicals generated within cytochrome c oxidase. These results suggest that ferricytochrome c may have an important role in protection of cytochrome c oxidase and consequently the mitochondrion against oxidative damage.
Published by Elsevier Inc.
Figures
References
-
- Chance B, Williams GR. The respiratory chain and oxidative phosphorylation. Advances in Enzymol. 1956;17:65–134. - PubMed
-
- Lesnefsky EJ, Hoppel CL. Ischemia-reperfusion injury in the aged heart: role of mitochondria. Arch. Biochem. Biophys. 2003;420:287–297. - PubMed
-
- Leeuwenburgh C, Rasmussen JE, Hsu FF, Mueller DM, Pennathur S, Heinecke JW. Mass spectrometric quantification of markers for protein oxidation by tyrosyl radical, cooper, and hydroxyl radical in low density lipoprotein isolated from human atherosclerotic plaques. J. Biol. Chem. 1997;272:3520–3526. - PubMed
-
- Cohen G. Oxidative stress, mitochondrial respiration, and Parkinson’s disease. Ann. N.Y. Acad. Sci. 2000;899:112–120. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
