

RPL30 regulation of splicing reveals distinct roles for Cbp80 in U1 and U2 snRNP cotranscriptional recruitment
- PMID: 20801768
- PMCID: PMC2941111
- DOI: 10.1261/rna.2366310
RPL30 regulation of splicing reveals distinct roles for Cbp80 in U1 and U2 snRNP cotranscriptional recruitment
Abstract
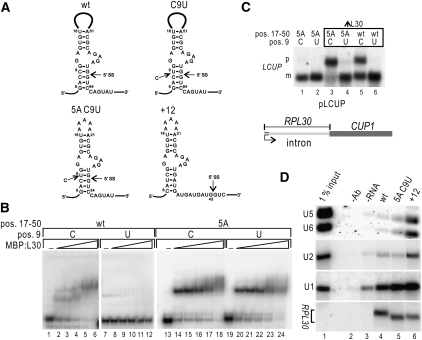
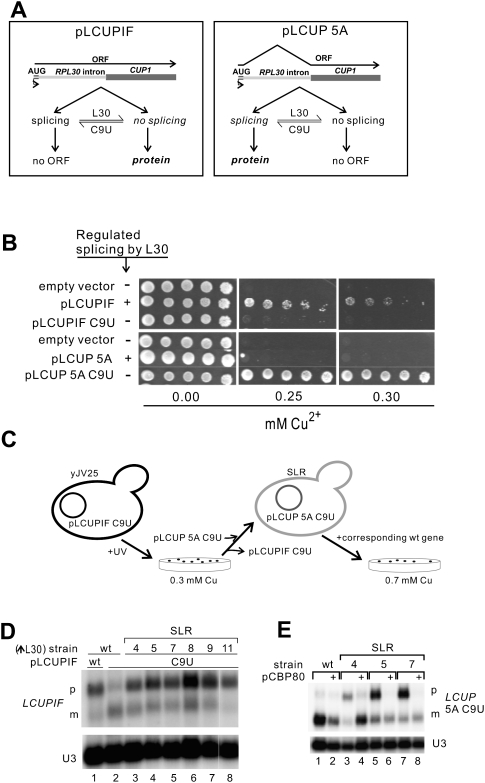
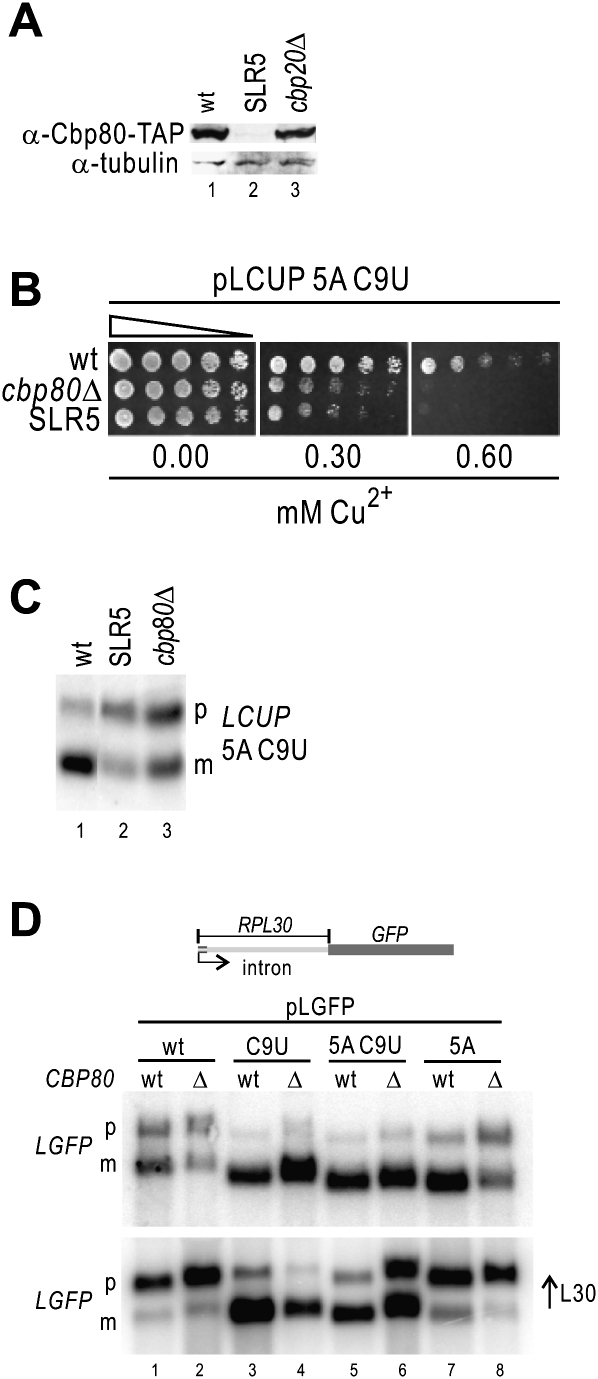
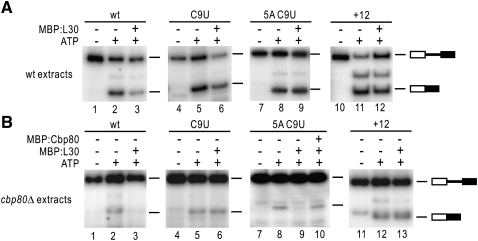
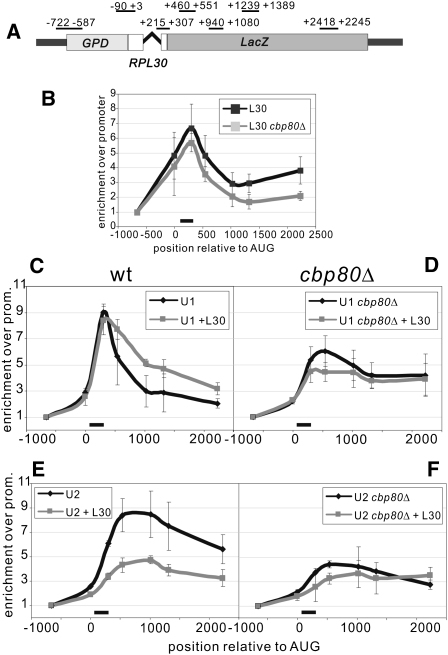
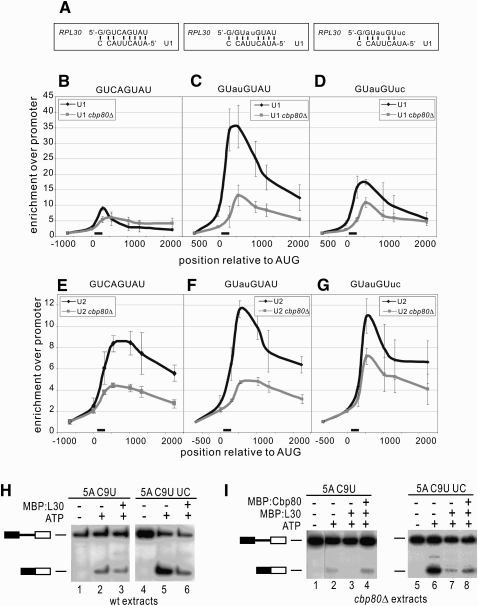
Pre-mRNA splicing is catalyzed by the spliceosome, and its control is essential for correct gene expression. While splicing repressors typically interfere with transcript recognition by spliceosomal components, the yeast protein L30 blocks spliceosomal rearrangements required for the engagement of U2 snRNP (small ribonucleoprotein particle) to its own transcript RPL30. Using a mutation in the RPL30 binding site that disrupts this repression, we have taken a genetic approach to reveal that regulation of splicing is restored in this mutant by deletion of the cap-binding complex (CBC) component Cbp80. Indeed, our data indicate that Cbp80 plays distinct roles in the recognition of the intron by U1 and U2 snRNP. It promotes the initial 5' splice site recognition by U1 and, independently, facilitates U2 recruitment, depending on sequences located in the vicinity of the 5' splice site. These results reveal a novel function for CBC in splicing and imply that these molecular events can be the target of a splicing regulator.
Figures
References
-
- Abovich N, Rosbash M 1997. Cross-intron bridging interactions in the yeast commitment complex are conserved in mammals. Cell 89: 403–412 - PubMed
-
- Abovich N, Liao XC, Rosbash M 1994. The yeast MUD2 protein: An interaction with PRP11 defines a bridge between commitment complexes and U2 snRNP addition. Genes Dev 8: 843–854 - PubMed
-
- Berget SM 1995. Exon recognition in vertebrate splicing. J Biol Chem 270: 2411–2414 - PubMed
-
- Brow DA 2002. Allosteric cascade of spliceosome activation. Annu Rev Genet 36: 333–360 - PubMed
-
- Chao JA, Williamson JR 2004. Joint X-ray and NMR refinement of the yeast L30e-mRNA complex. Structure 12: 1165–1176 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous