

Aberrant overexpression of the Rgl2 Ral small GTPase-specific guanine nucleotide exchange factor promotes pancreatic cancer growth through Ral-dependent and Ral-independent mechanisms
- PMID: 20801877
- PMCID: PMC2966088
- DOI: 10.1074/jbc.M110.116756
Aberrant overexpression of the Rgl2 Ral small GTPase-specific guanine nucleotide exchange factor promotes pancreatic cancer growth through Ral-dependent and Ral-independent mechanisms
Abstract
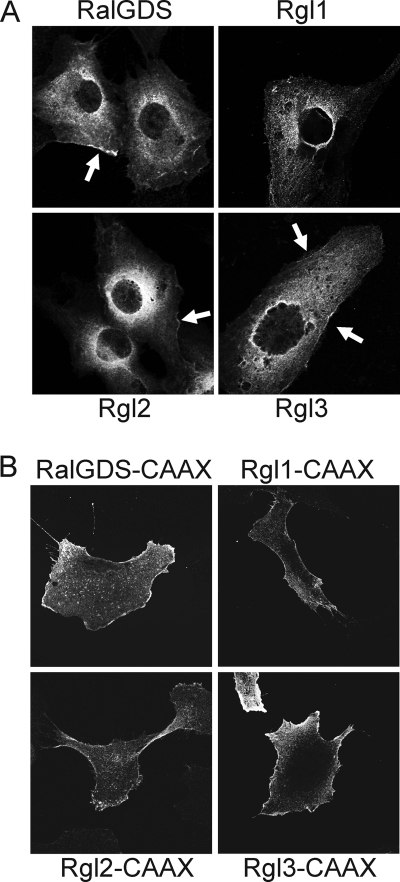
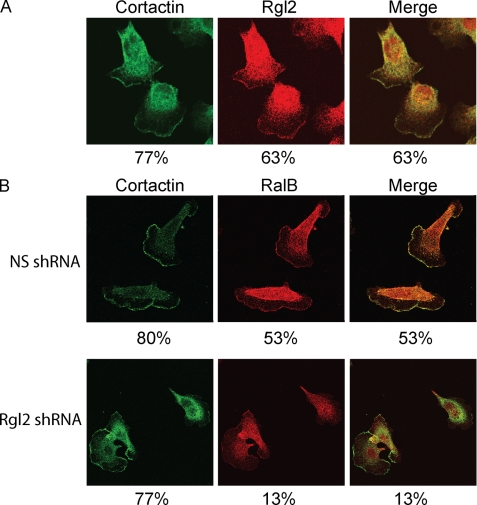
Our recent studies established essential and distinct roles for RalA and RalB small GTPase activation in K-Ras mutant pancreatic ductal adenocarcinoma (PDAC) cell line tumorigencity, invasion, and metastasis. However, the mechanism of Ral GTPase activation in PDAC has not been determined. There are four highly related mammalian RalGEFs (RalGDS, Rgl1, Rgl2, and Rgl3) that can serve as Ras effectors. Whether or not they share distinct or overlapping functions in K-Ras-mediated growth transformation has not been explored. We found that plasma membrane targeting to mimic persistent Ras activation enhanced the growth-transforming activities of RalGEFs. Unexpectedly, transforming activity did not correlate directly with total cell steady-state levels of Ral activation. Next, we observed elevated Rgl2 expression in PDAC tumor tissue and cell lines. Expression of dominant negative Ral, which blocks RalGEF function, as well as interfering RNA suppression of Rgl2, reduced PDAC cell line steady-state Ral activity, growth in soft agar, and Matrigel invasion. Surprisingly, the effect of Rgl2 on anchorage-independent growth could not be rescued by constitutively activated RalA, suggesting a novel Ral-independent function for Rgl2 in transformation. Finally, we determined that Rgl2 and RalB both localized to the leading edge, and this localization of RalB was dependent on endogenous Rgl2 expression. In summary, our observations support nonredundant roles for RalGEFs in Ras-mediated oncogenesis and a key role for Rgl2 in Ral activation and Ral-independent PDAC growth.
Figures






Similar articles
-
Ral GTPase-activating protein regulates the malignancy of pancreatic ductal adenocarcinoma.Cancer Sci. 2021 Aug;112(8):3064-3073. doi: 10.1111/cas.14970. Epub 2021 Jun 15. Cancer Sci. 2021. PMID: 34009715 Free PMC article.
-
Divergent roles for RalA and RalB in malignant growth of human pancreatic carcinoma cells.Curr Biol. 2006 Dec 19;16(24):2385-94. doi: 10.1016/j.cub.2006.10.023. Curr Biol. 2006. PMID: 17174914
-
The RalB small GTPase mediates formation of invadopodia through a GTPase-activating protein-independent function of the RalBP1/RLIP76 effector.Mol Cell Biol. 2012 Apr;32(8):1374-86. doi: 10.1128/MCB.06291-11. Epub 2012 Feb 13. Mol Cell Biol. 2012. PMID: 22331470 Free PMC article.
-
Ral small GTPase signaling and oncogenesis: More than just 15minutes of fame.Biochim Biophys Acta. 2014 Dec;1843(12):2976-2988. doi: 10.1016/j.bbamcr.2014.09.004. Epub 2014 Sep 16. Biochim Biophys Acta. 2014. PMID: 25219551 Free PMC article. Review.
-
The RalGEF/Ral pathway: evaluating an intervention opportunity for Ras cancers.Enzymes. 2013;34 Pt. B:137-56. doi: 10.1016/B978-0-12-420146-0.00006-8. Epub 2013 Nov 7. Enzymes. 2013. PMID: 25034103 Review.
Cited by
-
The RAL Enigma: Distinct Roles of RALA and RALB in Cancer.Cells. 2022 May 14;11(10):1645. doi: 10.3390/cells11101645. Cells. 2022. PMID: 35626682 Free PMC article. Review.
-
Drivers of Gene Expression Dysregulation in Pancreatic Cancer.Trends Cancer. 2021 Jul;7(7):594-605. doi: 10.1016/j.trecan.2021.01.008. Epub 2021 Feb 19. Trends Cancer. 2021. PMID: 33618999 Free PMC article. Review.
-
Extracellular superoxide dismutase regulates the expression of small gtpase regulatory proteins GEFs, GAPs, and GDI.PLoS One. 2015 Mar 9;10(3):e0121441. doi: 10.1371/journal.pone.0121441. eCollection 2015. PLoS One. 2015. PMID: 25751262 Free PMC article.
-
Ral GTPase is essential for actin dynamics and Golgi apparatus distribution in mouse oocyte maturation.Cell Div. 2021 Jun 10;16(1):3. doi: 10.1186/s13008-021-00071-y. Cell Div. 2021. PMID: 34112192 Free PMC article.
-
RalGPS2 Is Essential for Survival and Cell Cycle Progression of Lung Cancer Cells Independently of Its Established Substrates Ral GTPases.PLoS One. 2016 May 5;11(5):e0154840. doi: 10.1371/journal.pone.0154840. eCollection 2016. PLoS One. 2016. PMID: 27149377 Free PMC article.
References
-
- Jones S., Zhang X., Parsons D. W., Lin J. C., Leary R. J., Angenendt P., Mankoo P., Carter H., Kamiyama H., Jimeno A., Hong S. M., Fu B., Lin M. T., Calhoun E. S., Kamiyama M., Walter K., Nikolskaya T., Nikolsky Y., Hartigan J., Smith D. R., Hidalgo M., Leach S. D., Klein A. P., Jaffee E. M., Goggins M., Maitra A., Iacobuzio-Donahue C., Eshleman J. R., Kern S. E., Hruban R. H., Karchin R., Papadopoulos N., Parmigiani G., Vogelstein B., Velculescu V. E., Kinzler K. W. (2008) Science 321, 1801–1806 - PMC - PubMed
-
- Yeh J. J., Der C. J. (2007) Expert Opin. Ther. Targets 11, 673–694 - PubMed
-
- Wennerberg K., Rossman K. L., Der C. J. (2005) J. Cell Sci. 118, 843–846 - PubMed
-
- Bodemann B. O., White M. A. (2008) Nat. Rev. Cancer 8, 133–140 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
