

The basolateral amygdala and nucleus accumbens core mediate dissociable aspects of drug memory reconsolidation
- PMID: 20802017
- PMCID: PMC2948876
- DOI: 10.1101/lm.1757410
The basolateral amygdala and nucleus accumbens core mediate dissociable aspects of drug memory reconsolidation
Abstract
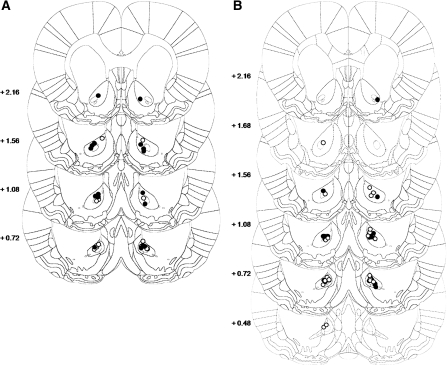
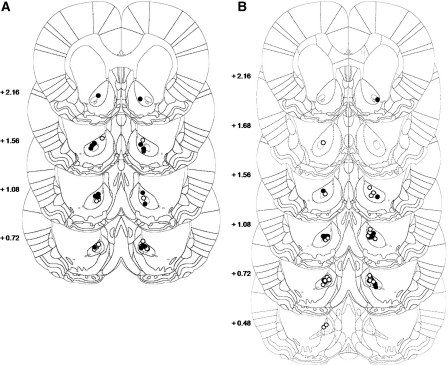
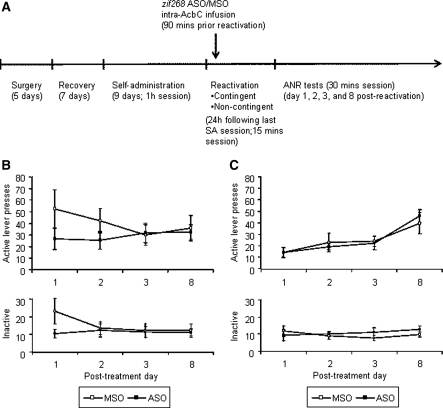
A distributed limbic-corticostriatal circuitry is implicated in cue-induced drug craving and relapse. Exposure to drug-paired cues not only precipitates relapse, but also triggers the reactivation and reconsolidation of the cue-drug memory. However, the limbic cortical-striatal circuitry underlying drug memory reconsolidation is unclear. The aim of this study was to investigate the involvement of the nucleus accumbens core and the basolateral amygdala in the reconsolidation of a cocaine-conditioned stimulus-evoked memory. Antisense oligodeoxynucleotides (ASO) were infused into each structure to knock down the expression of the immediate-early gene zif268, which is known to be required for memory reconsolidation. Control infusions used missense oligodeoxynucleotides (MSO). The effects of zif268 knockdown were measured in two complementary paradigms widely used to assess the impact of drug-paired CSs upon drug seeking: the acquisition of a new instrumental response with conditioned reinforcement and conditioned place preference. The results show that both intranucleus accumbens core and intrabasolateral amygdala zif268 ASO infusions at memory reactivation impaired the reconsolidation of the memory underlying a cocaine-conditioned place preference. However, knockdown of zif268 in the nucleus accumbens at memory reactivation had no effect on the memory underlying the conditioned reinforcing properties of the cocaine-paired CS measured subsequently, and this is in contrast to the marked impairment observed previously following intrabasolateral amygdala zif268 ASO infusions. These results suggest that both the basolateral amygdala and nucleus accumbens core are key structures within limbic cortical-striatal circuitry where reconsolidation of a cue-drug memory occurs. However reconsolidation of memory representations formed during Pavlovian conditioning are differentially localized in each site.
Figures




References
-
- Balleine BW 2005. Neural bases of food-seeking: Affect, arousal and reward in corticostriatolimbic circuits. Physiol Behav 86: 717–730 - PubMed
-
- Brunet A, Orr SP, Tremblay J, Robertson K, Nader K, Pitman RK 2008. Effect of post-retrieval propranolol on psychophysiologic responding during subsequent script-driven traumatic imagery in post-traumatic stress disorder. J Psychiatr Res 42: 503–506 - PubMed
-
- Cador M, Robbins TW, Everitt BJ 1989. Involvement of the amygdala in stimulus-reward associations: Interaction with the ventral striatum. Neuroscience 30: 77–86 - PubMed
-
- Cardinal RN, Aitken MRF 2006. ANOVA for the behavioural sciences researcher. Lawrence Erlbaum Associates, Mahwah, NJ
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources