

Biogenesis of Salmonella enterica serovar typhimurium membrane vesicles provoked by induction of PagC
- PMID: 20802043
- PMCID: PMC2953678
- DOI: 10.1128/JB.00590-10
Biogenesis of Salmonella enterica serovar typhimurium membrane vesicles provoked by induction of PagC
Abstract
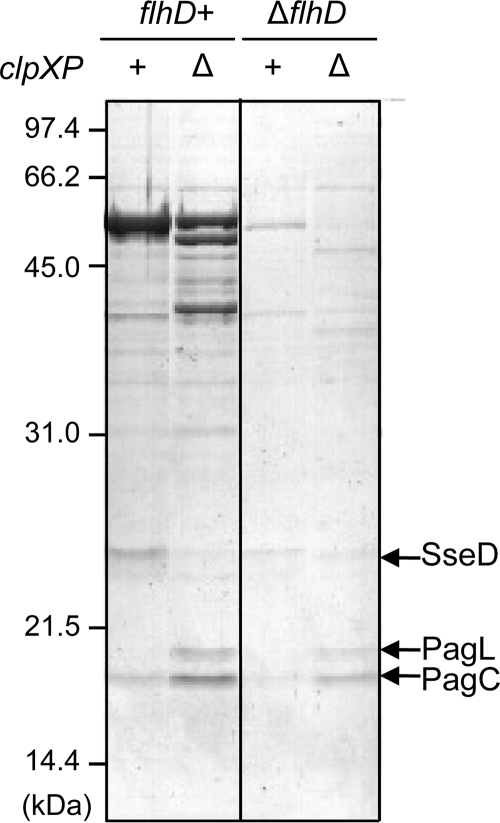
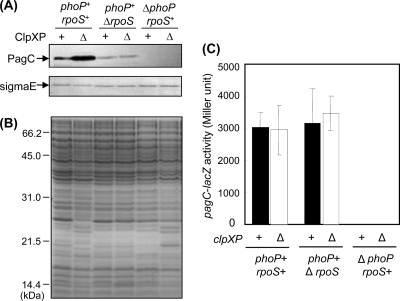
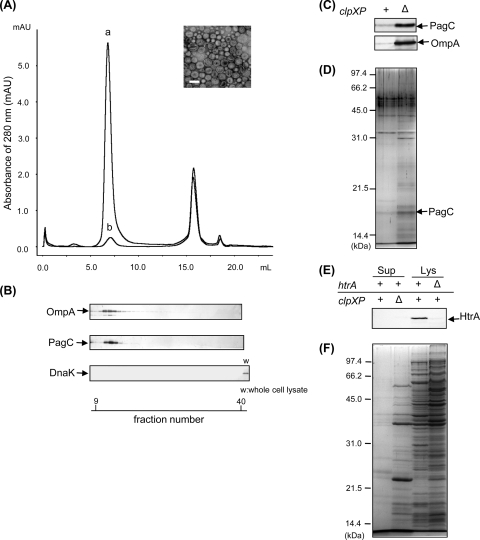
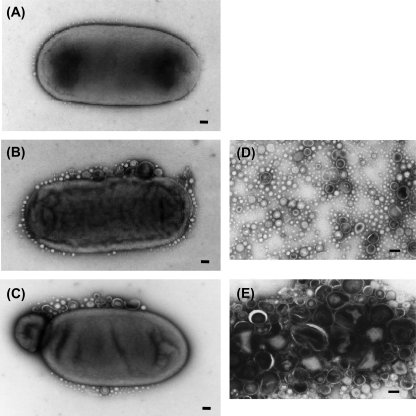
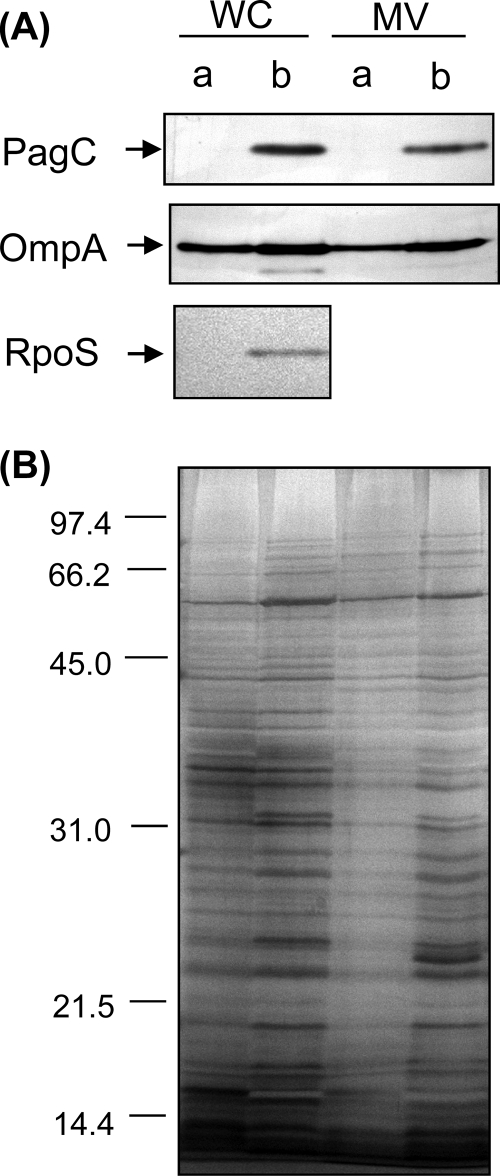
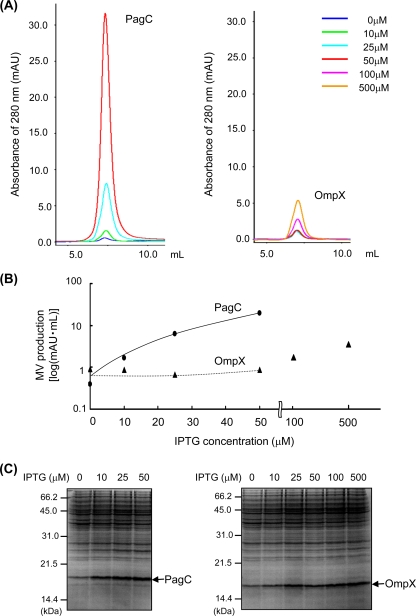
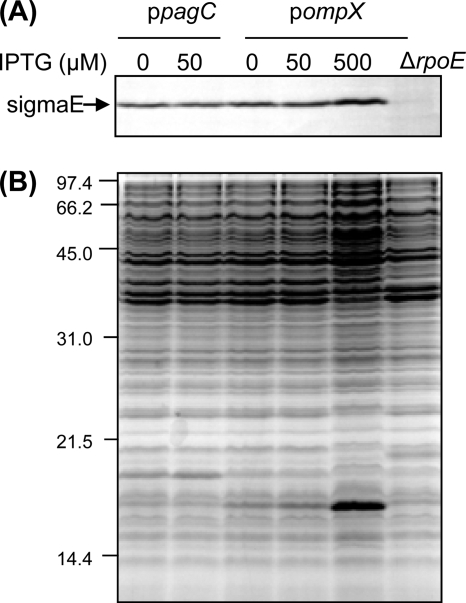
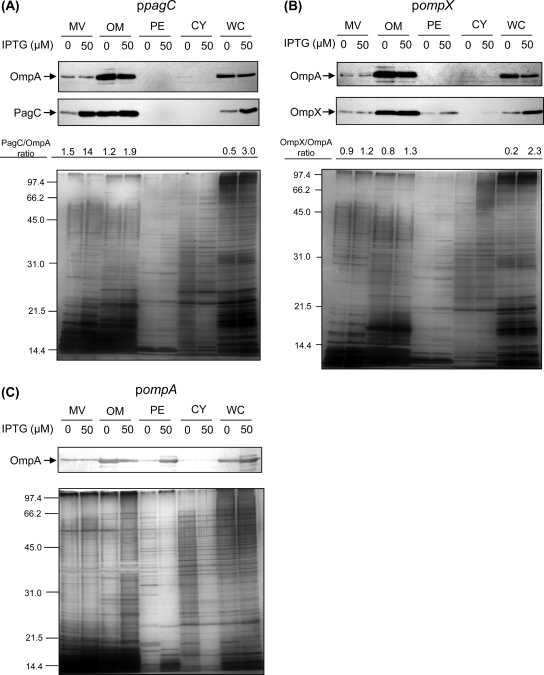
Gram-negative bacteria ubiquitously release membrane vesicles (MVs) into the extracellular milieu. Although MVs are the product of growing bacteria, not of cell lysis or death, the regulatory mechanisms underlying MV formation remained unknown. We have found that MV biogenesis is provoked by the induction of PagC, a Salmonella-specific protein whose expression is activated by conditions that mimic acidified macrophage phagosomes. PagC is a major constituent of Salmonella MVs, and increased expression accelerates vesiculation. Expression of PagC is regulated at the posttranscriptional and/or posttranslational level in a sigmaS (RpoS)-dependent manner. Serial quantitative analysis has demonstrated that MV formation can accelerate when the quantity of the MV constituents, OmpX and PagC, rises. Overproduction of PagC dramatically impacts the difference in the relative amount of vesiculation, but the corresponding overproduction of OmpX was less pronounced. Quantitative examination of the ratios of PagC and OmpX in the periplasm, outer membrane, and MVs demonstrates that PagC is preferentially enriched in MVs released from Salmonella cells. This suggests that specific protein sorting mechanisms operate when MVs are formed. The possible role(s) of PagC-MV in host cells is discussed.
Figures
References
-
- Alix, E., T. Miki, C. Felix, C. Rang, N. Figueroa-Bossi, E. Demettre, and A. B. Blanc-Potard. 2008. Interplay between MgtC and PagC in Salmonella enterica serovar Typhimurium. Microb. Pathog. 45:236-240. - PubMed
-
- Arnold, T., M. Poynor, S. Nussberger, A. N. Lupas, and D. Linke. 2007. Gene duplication of the eight-stranded β-barrel OmpX produces a functional pore: a scenario for the evolution of transmembrane β-barrels. J. Mol. Biol. 366:1174-1184. - PubMed
-
- Balsalobre, C., J. M. Silvan, S. Berglund, Y. Mizunoe, B. E. Uhlin, and S. N. Wai. 2006. Release of the type I secreted alpha-haemolysin via outer membrane vesicles from Escherichia coli. Mol. Microbiol. 59:99-112. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
