

RNA helicase A is a DNA-binding partner for EGFR-mediated transcriptional activation in the nucleus
- PMID: 20802156
- PMCID: PMC2941329
- DOI: 10.1073/pnas.1000743107
RNA helicase A is a DNA-binding partner for EGFR-mediated transcriptional activation in the nucleus
Abstract
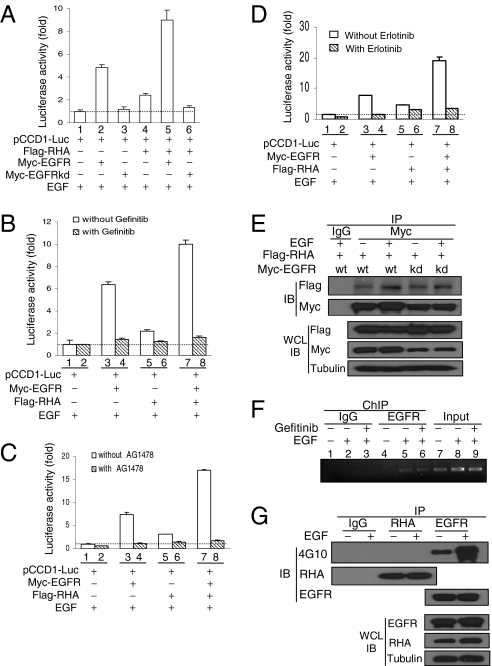
EGF induces the translocation of EGF receptor (EGFR) from the cell surface to the nucleus where EGFR activates gene transcription through its binding to an AT-rich sequence (ATRS) of the target gene promoter. However, how EGFR, without a DNA-binding domain, can bind to the gene promoter is unclear. In the present study, we show that RNA helicase A (RHA) is an important mediator for EGFR-induced gene transactivation. EGF stimulates the interaction of EGFR with RHA in the nucleus of cancer cells. The EGFR/RHA complex then associates with the target gene promoter through binding of RHA to the ATRS of the target gene promoter to activate its transcription. Knockdown of RHA expression in cancer cells abrogates the binding of EGFR to the target gene promoter, thereby reducing EGF/EGFR-induced gene expression. In addition, interruption of EGFR-RHA interaction decreases the EGFR-induced promoter activity. Consistently, we observed a positive correlation of the nuclear expression of EGFR, RHA, and cyclin D1 in human breast cancer samples. These results indicate that RHA is a DNA-binding partner for EGFR-mediated transcriptional activation in the nucleus.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Similar articles
-
PML represses lung cancer metastasis by suppressing the nuclear EGFR-mediated transcriptional activation of MMP2.Cell Cycle. 2014;13(19):3132-42. doi: 10.4161/15384101.2014.949212. Cell Cycle. 2014. PMID: 25486572 Free PMC article.
-
Sequence-specific DNA binding activity of RNA helicase A to the p16INK4a promoter.J Biol Chem. 2001 Jan 12;276(2):1634-42. doi: 10.1074/jbc.M004481200. J Biol Chem. 2001. PMID: 11038348
-
MUC1 regulates nuclear localization and function of the epidermal growth factor receptor.J Cell Sci. 2010 May 15;123(Pt 10):1716-23. doi: 10.1242/jcs.062661. Epub 2010 Apr 20. J Cell Sci. 2010. PMID: 20406885 Free PMC article.
-
Regulation of gene expression by progesterone in cancer cells: effects on cyclin D1, EGFR and VEGF.Mini Rev Med Chem. 2013 Apr 1;13(5):635-42. doi: 10.2174/1389557511313050002. Mini Rev Med Chem. 2013. PMID: 23432380 Review.
-
Nuclear EGFR signalling network in cancers: linking EGFR pathway to cell cycle progression, nitric oxide pathway and patient survival.Br J Cancer. 2006 Jan 30;94(2):184-8. doi: 10.1038/sj.bjc.6602941. Br J Cancer. 2006. PMID: 16434982 Free PMC article. Review.
Cited by
-
The DEAD-Box RNA Helicase DDX3 Interacts with NF-κB Subunit p65 and Suppresses p65-Mediated Transcription.PLoS One. 2016 Oct 13;11(10):e0164471. doi: 10.1371/journal.pone.0164471. eCollection 2016. PLoS One. 2016. PMID: 27736973 Free PMC article.
-
The Drosophila Helicase Maleless (MLE) is Implicated in Functions Distinct From its Role in Dosage Compensation.Mol Cell Proteomics. 2015 Jun;14(6):1478-88. doi: 10.1074/mcp.M114.040667. Epub 2015 Mar 16. Mol Cell Proteomics. 2015. PMID: 25776889 Free PMC article.
-
The translocon Sec61beta localized in the inner nuclear membrane transports membrane-embedded EGF receptor to the nucleus.J Biol Chem. 2010 Dec 3;285(49):38720-9. doi: 10.1074/jbc.M110.158659. Epub 2010 Oct 11. J Biol Chem. 2010. PMID: 20937808 Free PMC article.
-
Cellular functions of eukaryotic RNA helicases and their links to human diseases.Nat Rev Mol Cell Biol. 2023 Oct;24(10):749-769. doi: 10.1038/s41580-023-00628-5. Epub 2023 Jul 20. Nat Rev Mol Cell Biol. 2023. PMID: 37474727 Review.
-
Nuclear miR-150 enhances hepatic lipid accumulation by targeting RNA transcripts overlapping the PLIN2 promoter.iScience. 2023 Sep 7;26(10):107837. doi: 10.1016/j.isci.2023.107837. eCollection 2023 Oct 20. iScience. 2023. PMID: 37736048 Free PMC article.
References
-
- Linggi B, Carpenter G. ErbB receptors: New insights on mechanisms and biology. Trends Cell Biol. 2006;16:649–656. - PubMed
-
- Mosesson Y, Mills GB, Yarden Y. Derailed endocytosis: An emerging feature of cancer. Nat Rev Cancer. 2008;8:835–850. - PubMed
-
- Das AK, et al. Somatic mutations in the tyrosine kinase domain of epidermal growth factor receptor (EGFR) abrogate EGFR-mediated radioprotection in non-small cell lung carcinoma. Cancer Res. 2007;67:5267–5274. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
