

Heat shock transcription factor 1 localizes to sex chromatin during meiotic repression
- PMID: 20802198
- PMCID: PMC2966061
- DOI: 10.1074/jbc.M110.157552
Heat shock transcription factor 1 localizes to sex chromatin during meiotic repression
Abstract
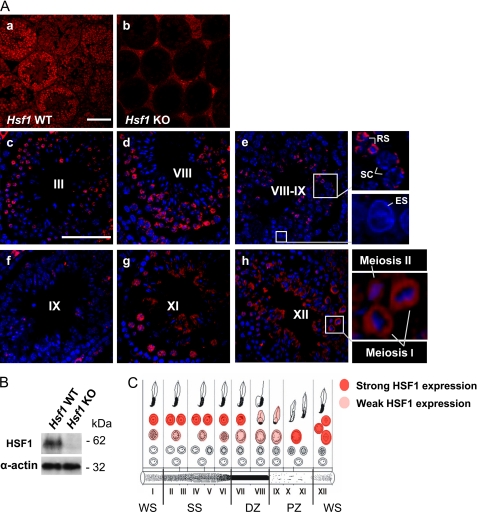
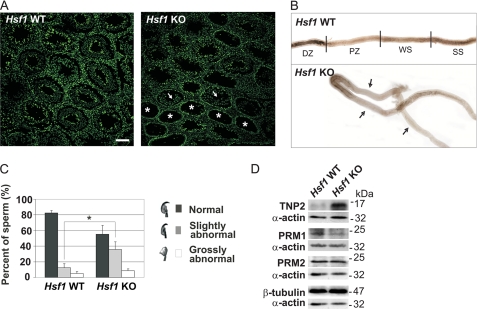
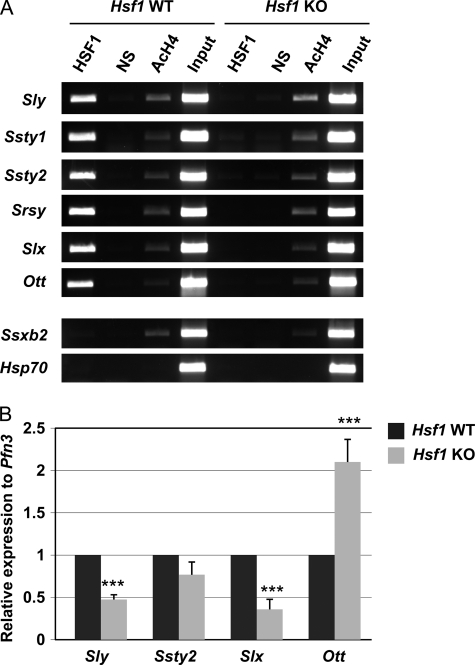
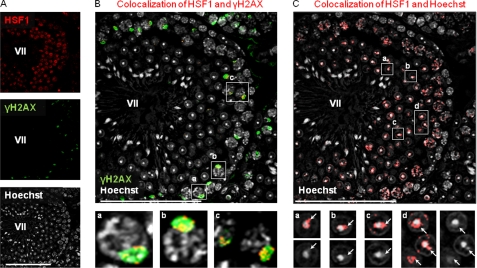
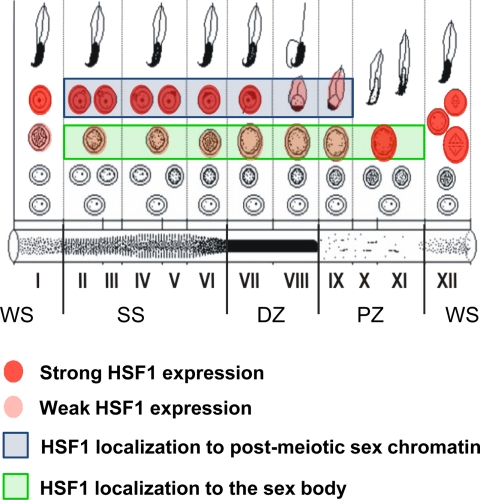
Heat shock factor 1 (HSF1) is an important transcription factor in cellular stress responses, cancer, aging, and developmental processes including gametogenesis. Disruption of Hsf1, together with another HSF family member, Hsf2, causes male sterility and complete lack of mature sperm in mice, but the specific role of HSF1 in spermatogenesis has remained unclear. Here, we show that HSF1 is transiently expressed in meiotic spermatocytes and haploid round spermatids in mouse testis. The Hsf1(-/-) male mice displayed regions of seminiferous tubules containing only spermatogonia and increased morphological abnormalities in sperm heads. In search for HSF1 target genes, we identified 742 putative promoters in mouse testis. Among them, the sex chromosomal multicopy genes that are expressed in postmeiotic cells were occupied by HSF1. Given that the sex chromatin mostly is repressed during and after meiosis, it is remarkable that HSF1 directly regulates the transcription of sex-linked multicopy genes during postmeiotic repression. In addition, our results show that HSF1 localizes to the sex body prior to the meiotic divisions and to the sex chromocenter after completed meiosis. To the best of our knowledge, HSF1 is the first known transcription factor found at the repressed sex chromatin during meiosis.
Figures
References
-
- Ellis P. J., Affara N. A. (2006) Hum. Fertil. (Camb.) 9, 1–7 - PubMed
-
- Hoyer-Fender S. (2003) Cytogenet. Genome Res. 103, 245–255 - PubMed
-
- Handel M. A. (2004) Exp. Cell. Res. 296, 57–63 - PubMed
-
- Turner J. M. (2007) Development 134, 1823–1831 - PubMed
-
- Solari A. J. (1974) Int. Rev. Cytol. 38, 273–317 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
