

ATR and ATM differently regulate WRN to prevent DSBs at stalled replication forks and promote replication fork recovery
- PMID: 20802463
- PMCID: PMC2944071
- DOI: 10.1038/emboj.2010.205
ATR and ATM differently regulate WRN to prevent DSBs at stalled replication forks and promote replication fork recovery
Abstract
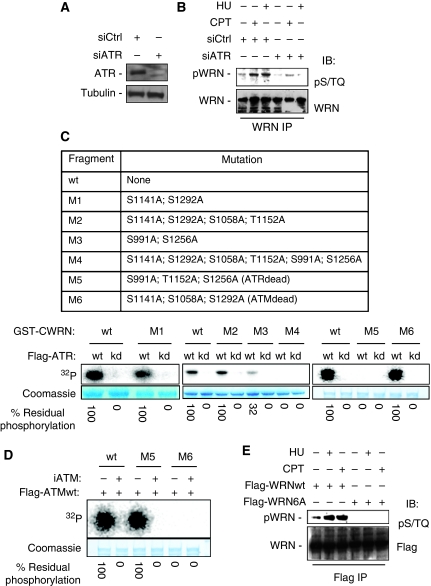
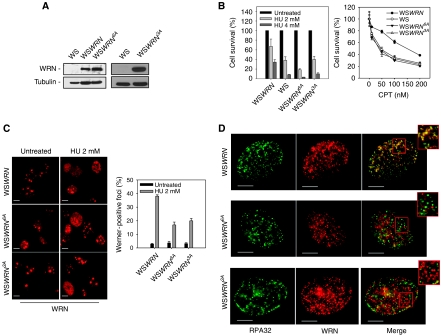
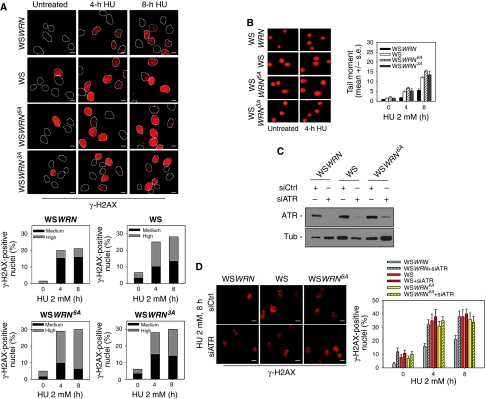
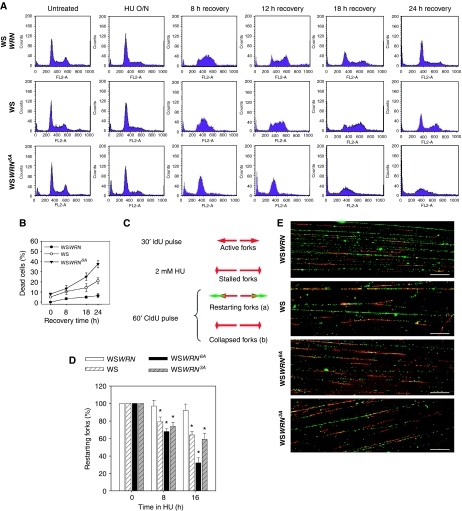
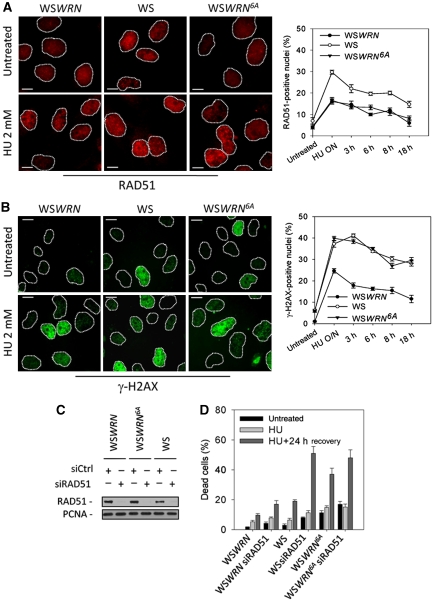
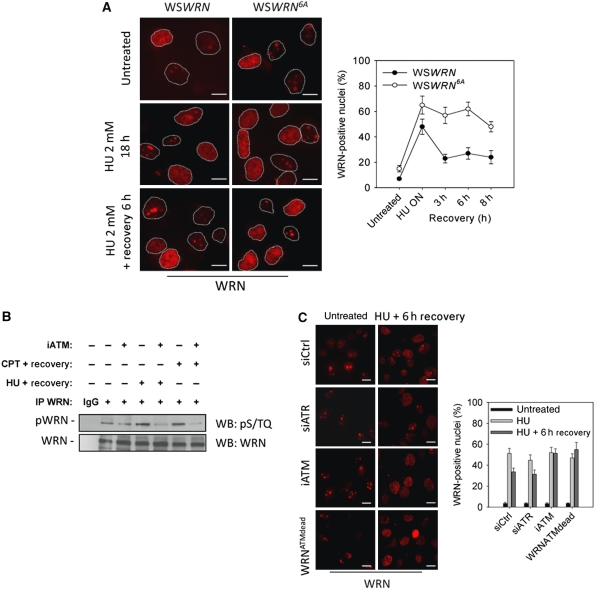
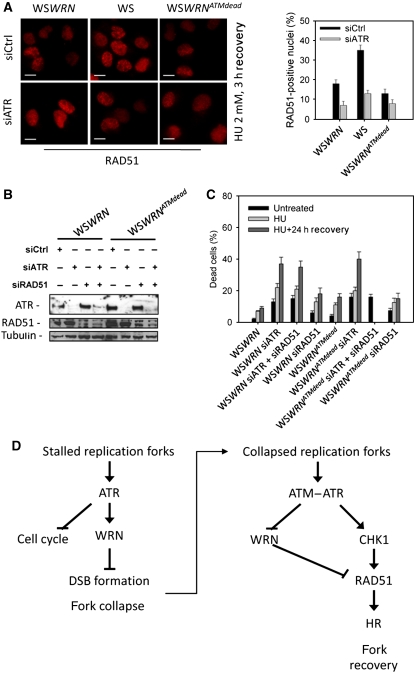
Accurate response to replication arrest is crucial to preserve genome stability and requires both the ATR and ATM functions. The Werner syndrome protein (WRN) is implicated in the recovery of stalled replication forks, and although an ATR/ATM-dependent phosphorylation of WRN was observed after replication arrest, the function of such modifications during the response to perturbed replication is not yet appreciated. Here, we report that WRN is directly phosphorylated by ATR at multiple C-terminal S/TQ residues. Suppression of ATR-mediated phosphorylation of WRN prevents proper accumulation of WRN in nuclear foci, co-localisation with RPA and causes breakage of stalled forks. On the other hand, inhibition of ATM kinase activity or expression of an ATM-unphosphorylable WRN allele leads to retention of WRN in nuclear foci and impaired recruitment of RAD51 recombinase resulting in reduced viability after fork collapse. Altogether, our findings indicate that ATR and ATM promote recovery from perturbed replication by differently regulating WRN at defined moments of the response to replication fork arrest.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Ababou M, Dutertre S, Lecluse Y, Onclercq R, Chatton B, Amor-Gueret M (2000) ATM-dependent phosphorylation and accumulation of endogenous BLM protein in response to ionizing radiation. Oncogene 19: 5955–5963 - PubMed
-
- Arlt MF, Durkin SG, Ragland RL, Glover TW (2006) Common fragile sites as targets for chromosome rearrangements. DNA Repair (Amst) 5: 1126–1135 - PubMed
-
- Arnaudeau C, Lundin C, Helleday T (2001) DNA double-strand breaks associated with replication forks are predominantly repaired by homologous recombination involving an exchange mechanism in mammalian cells. J Mol Biol 307: 1235–1245 - PubMed
-
- Beamish H, Kedar P, Kaneko H, Chen P, Fukao T, Peng C, Beresten S, Gueven N, Purdie D, Lees-Miller S, Ellis N, Kondo N, Lavin MF (2002) Functional link between BLM defective in Bloom's syndrome and the ataxia-telangiectasia-mutated protein, ATM. J Biol Chem 277: 30515–30523 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
