

Identification of a quality-control mechanism for mRNA 5'-end capping
- PMID: 20802481
- PMCID: PMC2948066
- DOI: 10.1038/nature09338
Identification of a quality-control mechanism for mRNA 5'-end capping
Abstract
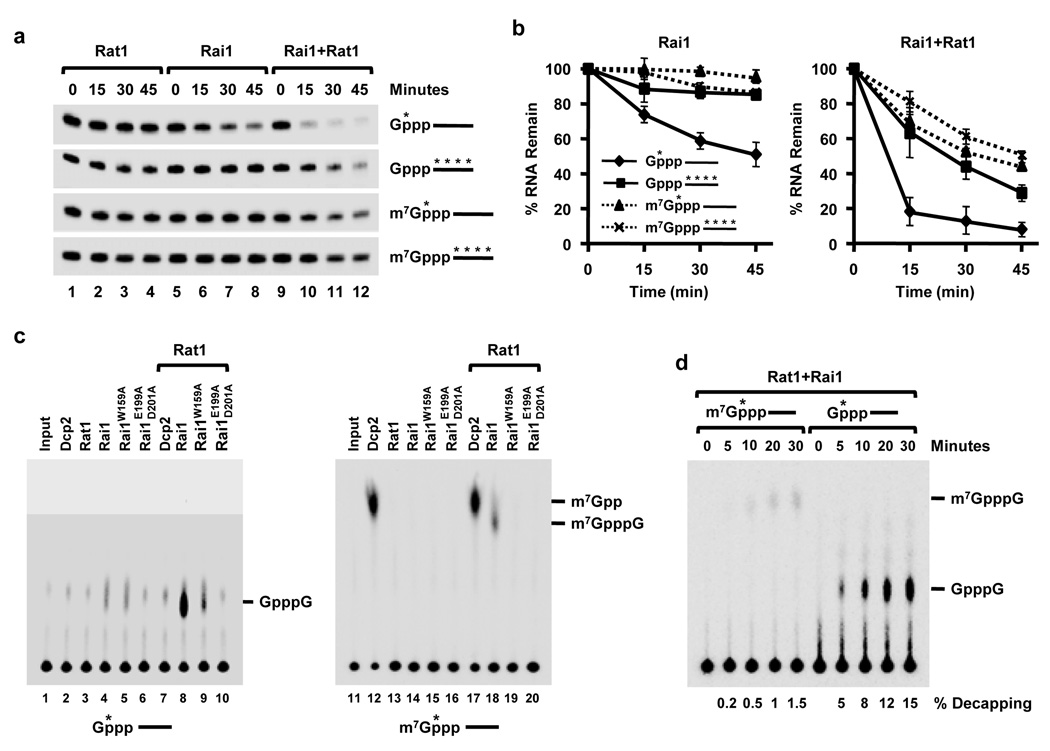
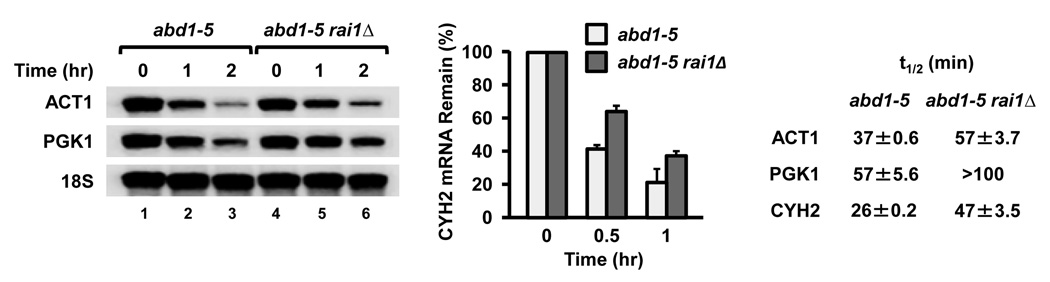
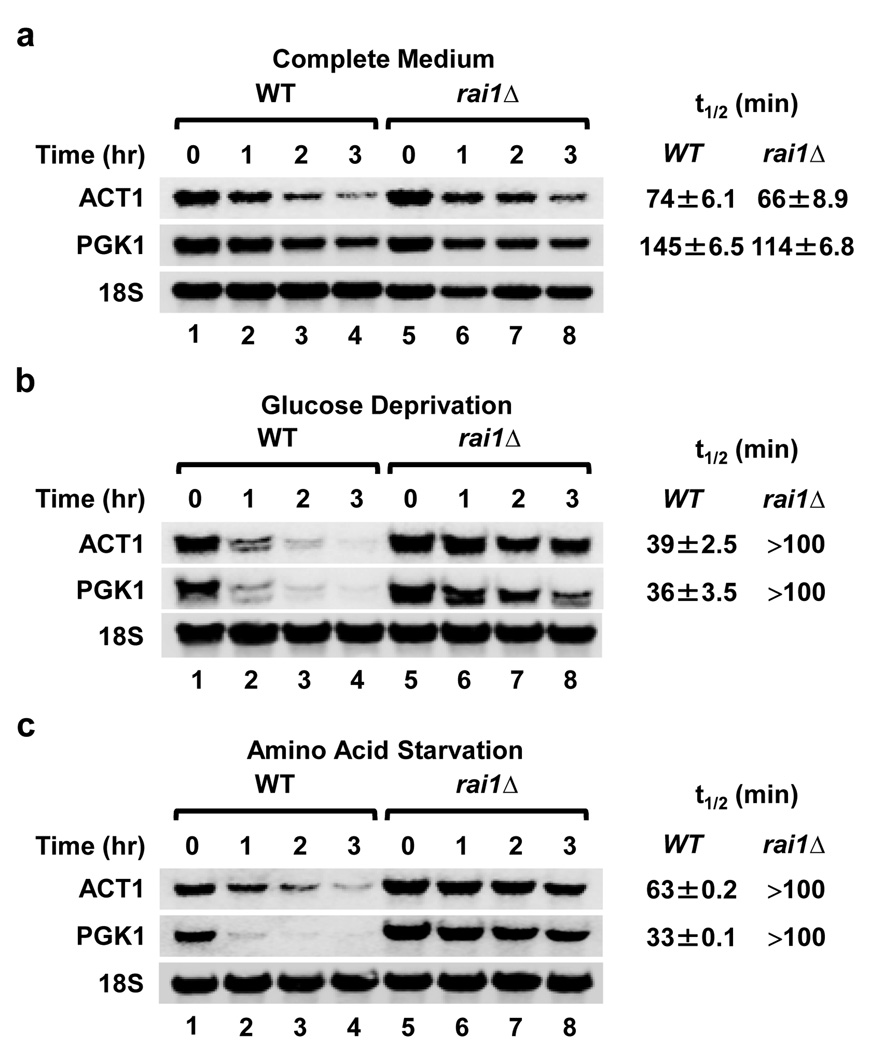
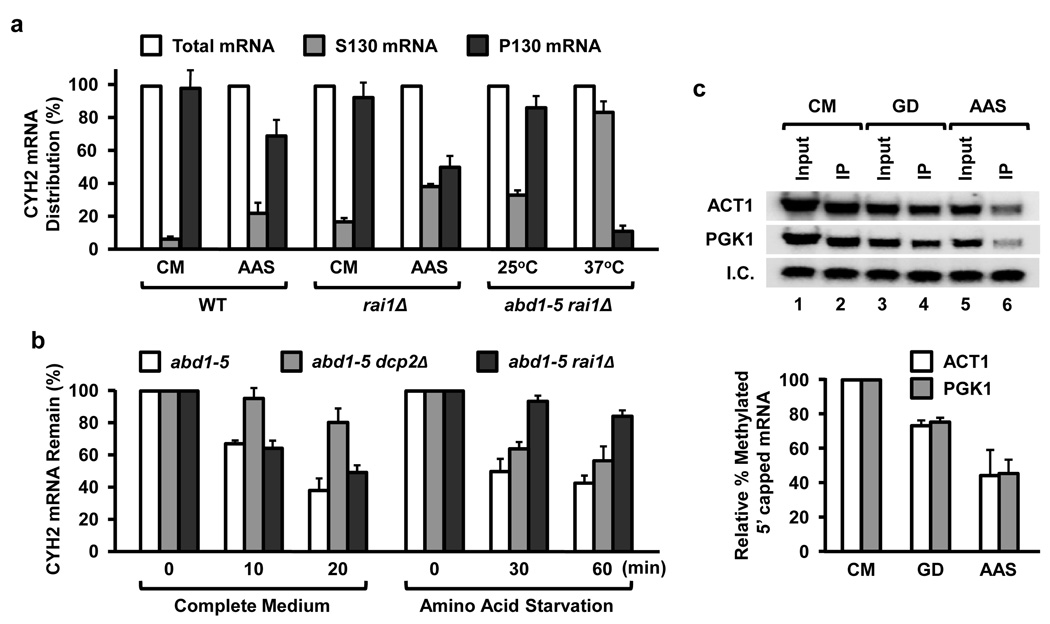
The 7-methylguanosine cap structure at the 5' end of eukaryotic messenger RNAs is a critical determinant of their stability and translational efficiency. It is generally believed that 5'-end capping is a constitutive process that occurs during mRNA maturation and lacks the need for a quality-control mechanism to ensure its fidelity. We recently reported that the yeast Rai1 protein has pyrophosphohydrolase activity towards mRNAs lacking a 5'-end cap. Here we show that, in vitro as well as in yeast cells, Rai1 possesses a novel decapping endonuclease activity that can also remove the entire cap structure dinucleotide from an mRNA. This activity is targeted preferentially towards mRNAs with unmethylated caps in contrast to the canonical decapping enzyme, Dcp2, which targets mRNAs with a methylated cap. Capped but unmethylated mRNAs generated in yeast cells with a defect in the methyltransferase gene are more stable in a rai1-gene-disrupted background. Moreover, rai1Δ yeast cells with wild-type capping enzymes show significant accumulation of mRNAs with 5'-end capping defects under nutritional stress conditions of glucose starvation or amino acid starvation. These findings provide evidence that 5'-end capping is not a constitutive process that necessarily always proceeds to completion and demonstrates that Rai1 has an essential role in clearing mRNAs with aberrant 5'-end caps. We propose that Rai1 is involved in an as yet uncharacterized quality control process that ensures mRNA 5'-end integrity by an aberrant-cap-mediated mRNA decay mechanism.
Figures
References
-
- Liu H, Kiledjian M. Decapping the message: a beginning or an end. Biochem Soc Trans. 2006;34:35–38. - PubMed
-
- Meyer S, Temme C, Wahle E. Messenger RNA turnover in eukaryotes: pathways and enzymes. Crit Rev Biochem Mol Biol. 2004;39:197–216. - PubMed
-
- Merrick WC. Cap-dependent and cap-independent translation in eukaryotic systems. Gene. 2004;332:1–11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
