

Hypothalamic AMPK and fatty acid metabolism mediate thyroid regulation of energy balance
- PMID: 20802499
- PMCID: PMC2935934
- DOI: 10.1038/nm.2207
Hypothalamic AMPK and fatty acid metabolism mediate thyroid regulation of energy balance
Abstract
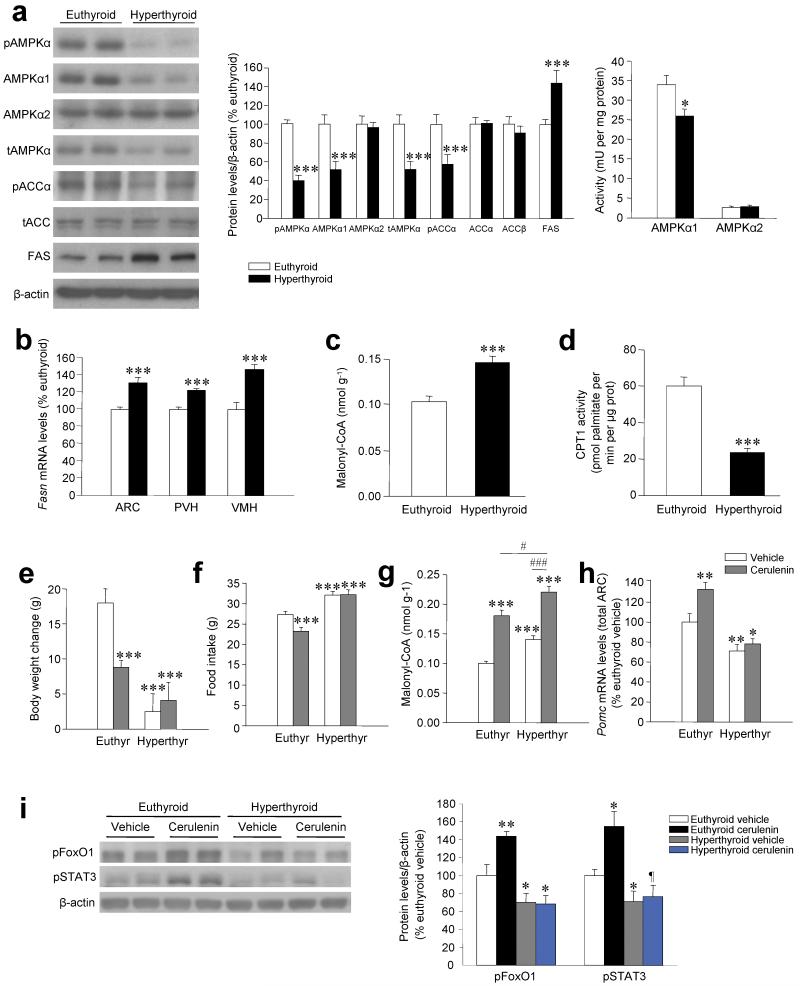
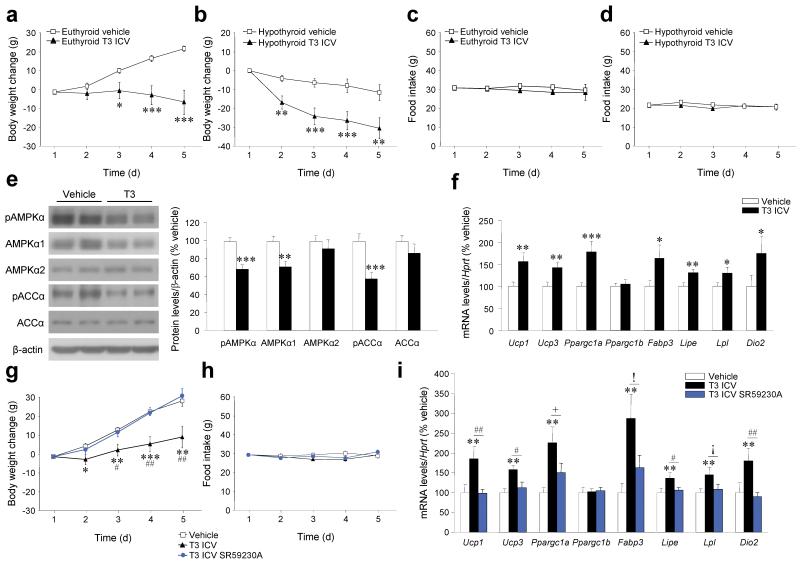
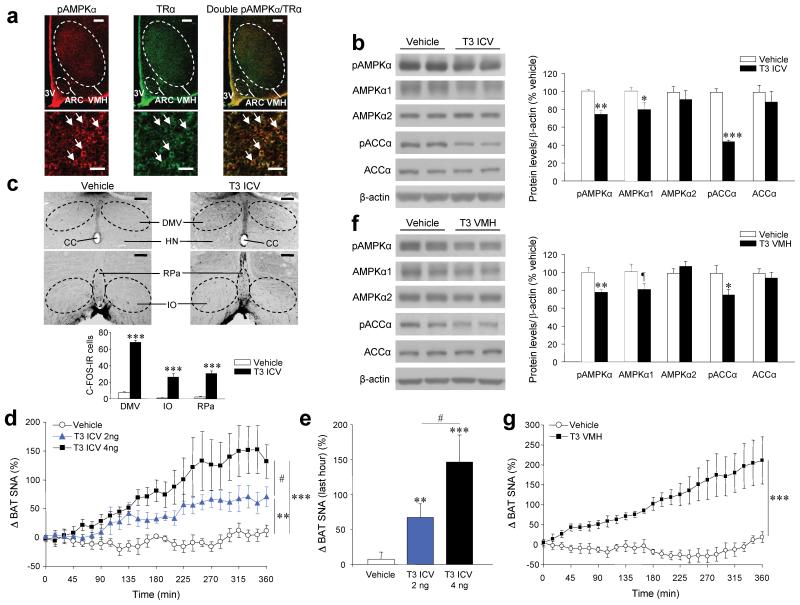
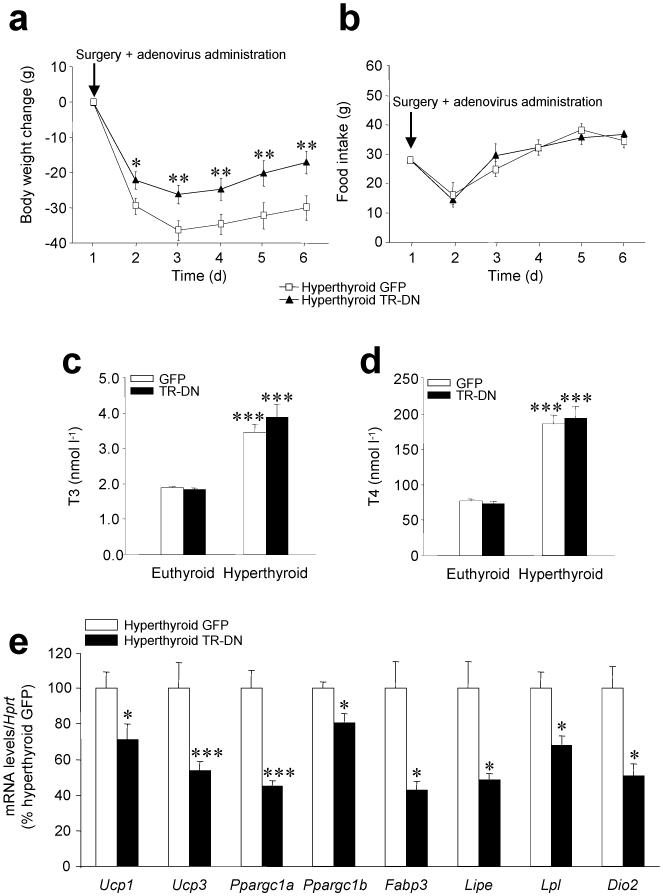
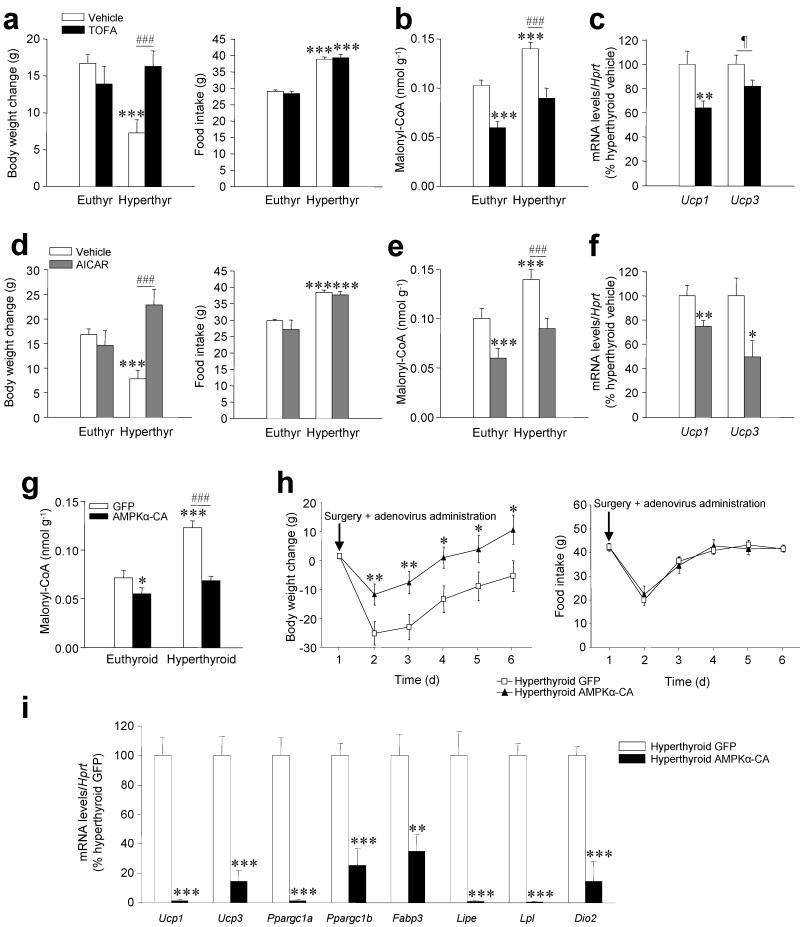
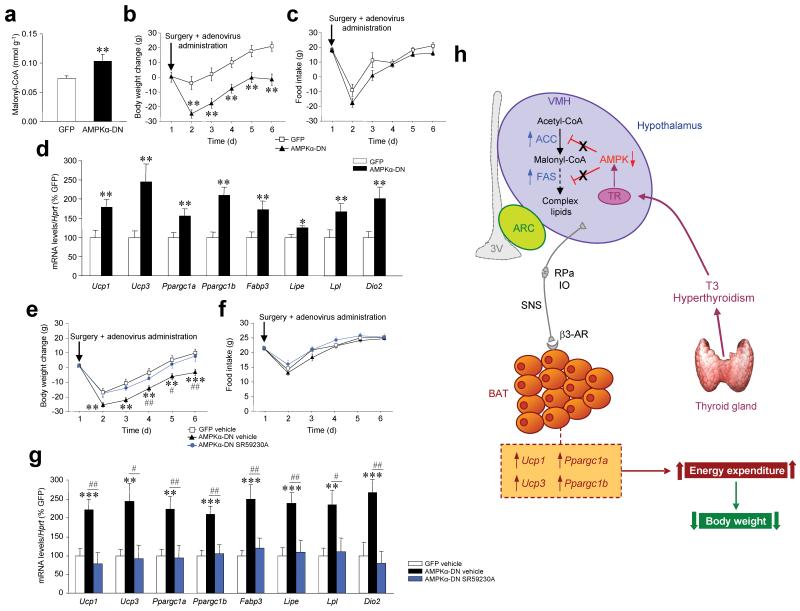
Thyroid hormones have widespread cellular effects; however it is unclear whether their effects on the central nervous system (CNS) contribute to global energy balance. Here we demonstrate that either whole-body hyperthyroidism or central administration of triiodothyronine (T3) decreases the activity of hypothalamic AMP-activated protein kinase (AMPK), increases sympathetic nervous system (SNS) activity and upregulates thermogenic markers in brown adipose tissue (BAT). Inhibition of the lipogenic pathway in the ventromedial nucleus of the hypothalamus (VMH) prevents CNS-mediated activation of BAT by thyroid hormone and reverses the weight loss associated with hyperthyroidism. Similarly, inhibition of thyroid hormone receptors in the VMH reverses the weight loss associated with hyperthyroidism. This regulatory mechanism depends on AMPK inactivation, as genetic inhibition of this enzyme in the VMH of euthyroid rats induces feeding-independent weight loss and increases expression of thermogenic markers in BAT. These effects are reversed by pharmacological blockade of the SNS. Thus, thyroid hormone-induced modulation of AMPK activity and lipid metabolism in the hypothalamus is a major regulator of whole-body energy homeostasis.
Figures
Comment in
-
Thyroid hormones: igniting brown fat via the brain.Nat Med. 2010 Sep;16(9):965-7. doi: 10.1038/nm0910-965. Nat Med. 2010. PMID: 20823876 No abstract available.
References
-
- Silva JE. Thyroid hormone control of thermogenesis and energy balance. Thyroid. 1995;5:481–492. - PubMed
-
- Pijl H, et al. Food choice in hyperthyroidism: potential influence of the autonomic nervous system and brain serotonin precursor availability. J. Clin. Endocrinol. Metab. 2001;86:5848–5853. - PubMed
-
- Cannon B, Nedergaard J. Brown adipose tissue: function and physiological significance. Physiol Rev. 2004;84:277–359. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
