

Hypoxia and cell cycle regulation of the von Hippel-Lindau tumor suppressor
- PMID: 20802534
- PMCID: PMC2995849
- DOI: 10.1038/onc.2010.395
Hypoxia and cell cycle regulation of the von Hippel-Lindau tumor suppressor
Abstract
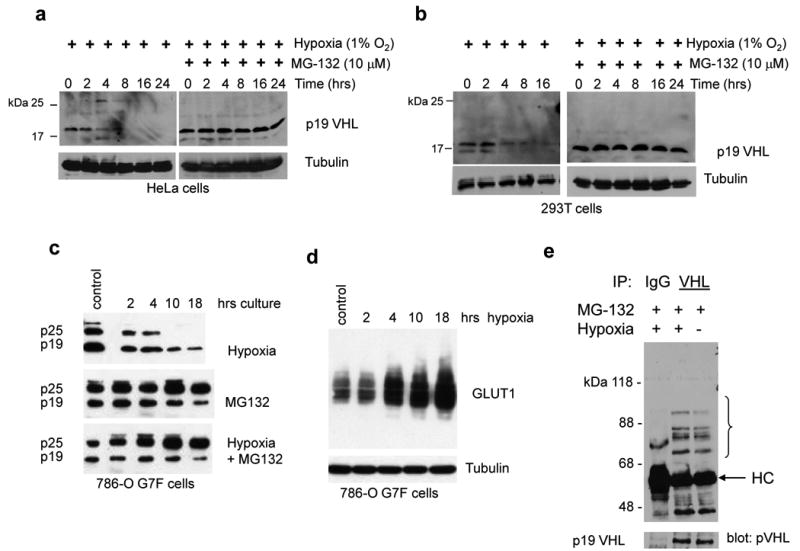
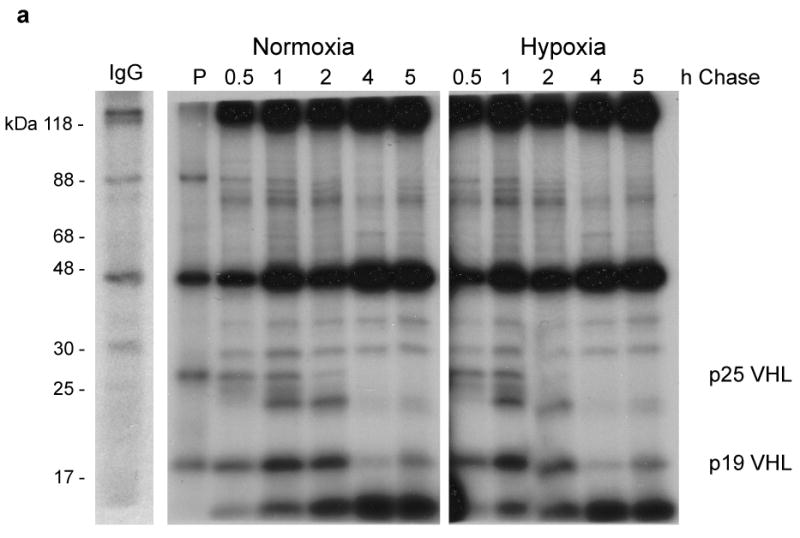
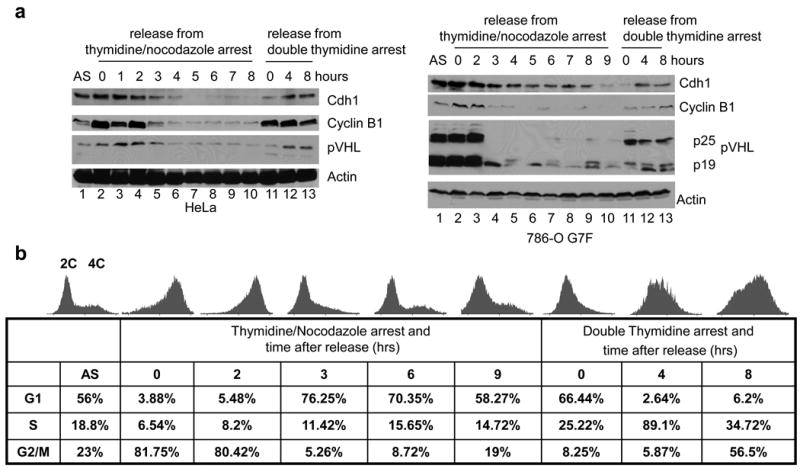
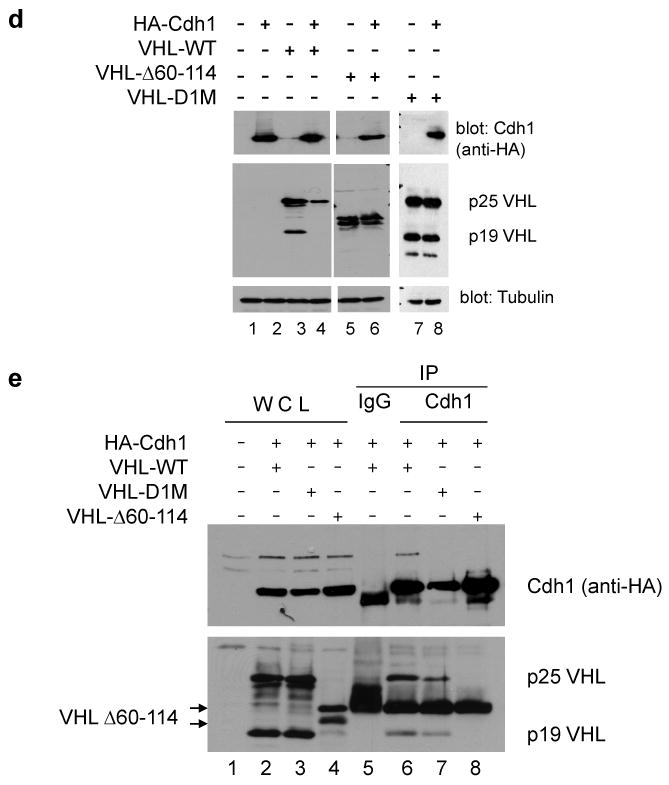
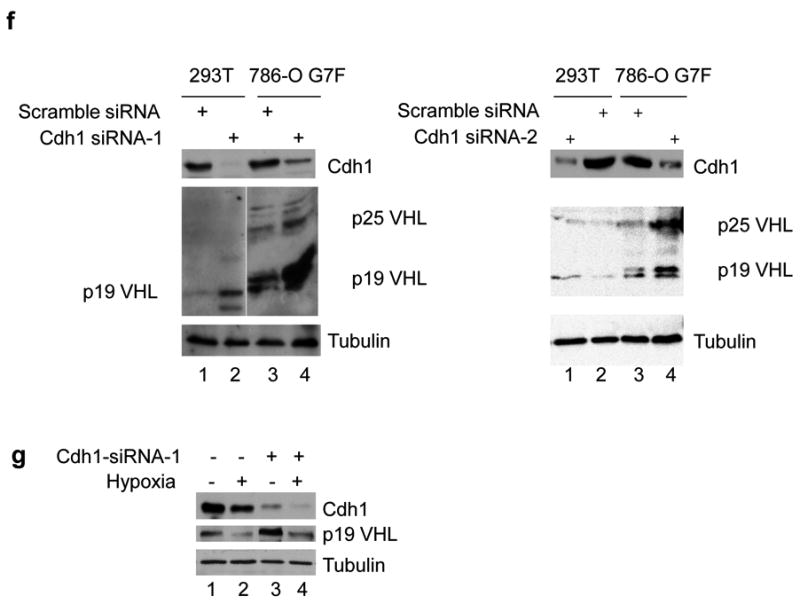
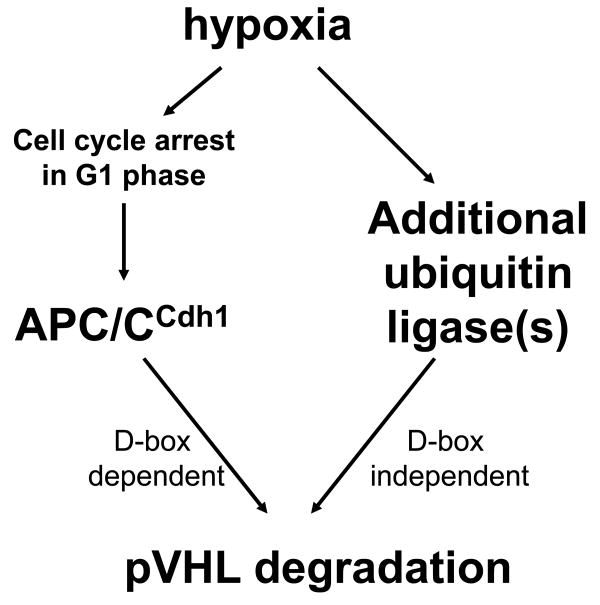
Inactivation of von Hippel-Lindau tumor-suppressor protein (pVHL) is associated with von Hippel-Lindau disease, an inherited cancer syndrome, as well as the majority of patients with sporadic clear cell renal cell carcinoma (RCC). Although the involvement of pVHL in oxygen sensing through targeting hypoxia-inducible factor-α subunits to ubiquitin-dependent proteolysis has been well documented, less is known about pVHL regulation under both normoxic and hypoxic conditions. We found that pVHL levels decreased in hypoxia and that hypoxia-induced cell cycle arrest is associated with pVHL expression in RCC cells. pVHL levels fluctuate during the cell cycle, paralleling cyclin B1 levels, with decreased levels in mitosis and G1. pVHL contains consensus destruction (D) box sequences, and pVHL associates with Cdh1, an activator of the anaphase-promoting complex/cyclosome (APC/C) E3 ubiquitin ligase. We show that pVHL has a decreased half-life in G1, Cdh1 downregulation results in increased pVHL expression, whereas Cdh1 overexpression results in decreased pVHL expression. Taken together, these results suggest that pVHL is a novel substrate of APC/C(Cdh1). D box-independent pVHL degradation was also detected, indicating that other ubiquitin ligases are also activated for pVHL degradation.
Figures






Similar articles
-
Identification of cyclin D1 and other novel targets for the von Hippel-Lindau tumor suppressor gene by expression array analysis and investigation of cyclin D1 genotype as a modifier in von Hippel-Lindau disease.Cancer Res. 2002 Jul 1;62(13):3803-11. Cancer Res. 2002. PMID: 12097293
-
[Von-Hippel-Lindau (VHL) protein function by initiation and progression of renal cancer].Pathologe. 2008 Nov;29 Suppl 2:149-52. doi: 10.1007/s00292-008-1034-y. Pathologe. 2008. PMID: 18751708 Review. German.
-
Contrasting effects on HIF-1alpha regulation by disease-causing pVHL mutations correlate with patterns of tumourigenesis in von Hippel-Lindau disease.Hum Mol Genet. 2001 May 1;10(10):1029-38. doi: 10.1093/hmg/10.10.1029. Hum Mol Genet. 2001. PMID: 11331613
-
The VHL tumor suppressor and HIF: insights from genetic studies in mice.Cell Death Differ. 2008 Apr;15(4):650-9. doi: 10.1038/sj.cdd.4402313. Epub 2008 Jan 25. Cell Death Differ. 2008. PMID: 18219317 Free PMC article. Review.
-
The role of von Hippel-Lindau tumor suppressor protein and hypoxia in renal clear cell carcinoma.Am J Physiol Renal Physiol. 2004 Jul;287(1):F1-6. doi: 10.1152/ajprenal.00424.2003. Am J Physiol Renal Physiol. 2004. PMID: 15180922 Review.
Cited by
-
Nitric oxide signaling in hypoxia.J Mol Med (Berl). 2012 Mar;90(3):217-31. doi: 10.1007/s00109-012-0880-5. Epub 2012 Feb 18. J Mol Med (Berl). 2012. PMID: 22349396 Review.
-
Novel interactions of the von Hippel-Lindau (pVHL) tumor suppressor with the CDKN1 family of cell cycle inhibitors.Sci Rep. 2017 Apr 20;7:46562. doi: 10.1038/srep46562. Sci Rep. 2017. PMID: 28425505 Free PMC article.
-
HIF-1α is over-expressed in leukemic cells from TP53-disrupted patients and is a promising therapeutic target in chronic lymphocytic leukemia.Haematologica. 2020 Apr;105(4):1042-1054. doi: 10.3324/haematol.2019.217430. Epub 2019 Jul 9. Haematologica. 2020. PMID: 31289209 Free PMC article.
-
Association of the von Hippel-Lindau protein with AUF1 and posttranscriptional regulation of VEGFA mRNA.Mol Cancer Res. 2012 Jan;10(1):108-20. doi: 10.1158/1541-7786.MCR-11-0435. Epub 2011 Nov 15. Mol Cancer Res. 2012. PMID: 22086907 Free PMC article.
-
RHOBTB3 promotes proteasomal degradation of HIFα through facilitating hydroxylation and suppresses the Warburg effect.Cell Res. 2015 Sep;25(9):1025-42. doi: 10.1038/cr.2015.90. Epub 2015 Jul 28. Cell Res. 2015. PMID: 26215701 Free PMC article.
References
-
- Beroud C, Collod-Beroud G, Boileau C, Soussi T, Junien C. UMD (Universal mutation database): a generic software to build and analyze locus-specific databases. Hum Mutat. 2000;15:86–94. - PubMed
-
- de Paulsen N, Brychzy A, Fournier MC, Klausner RD, Gnarra JR, Pause A, et al. Role of transforming growth factor-alpha in von Hippel--Lindau (VHL)(-/-) clear cell renal carcinoma cell proliferation: a possible mechanism coupling VHL tumor suppressor inactivation and tumorigenesis. Proc Natl Acad Sci U S A. 2001;98:1387–92. - PMC - PubMed
-
- Deshaies RJ, Joazeiro CA. RING domain E3 ubiquitin ligases. Annu Rev Biochem. 2009;78:399–434. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
