

PI(3,5)P(2) controls membrane trafficking by direct activation of mucolipin Ca(2+) release channels in the endolysosome
- PMID: 20802798
- PMCID: PMC2928581
- DOI: 10.1038/ncomms1037
PI(3,5)P(2) controls membrane trafficking by direct activation of mucolipin Ca(2+) release channels in the endolysosome
Abstract
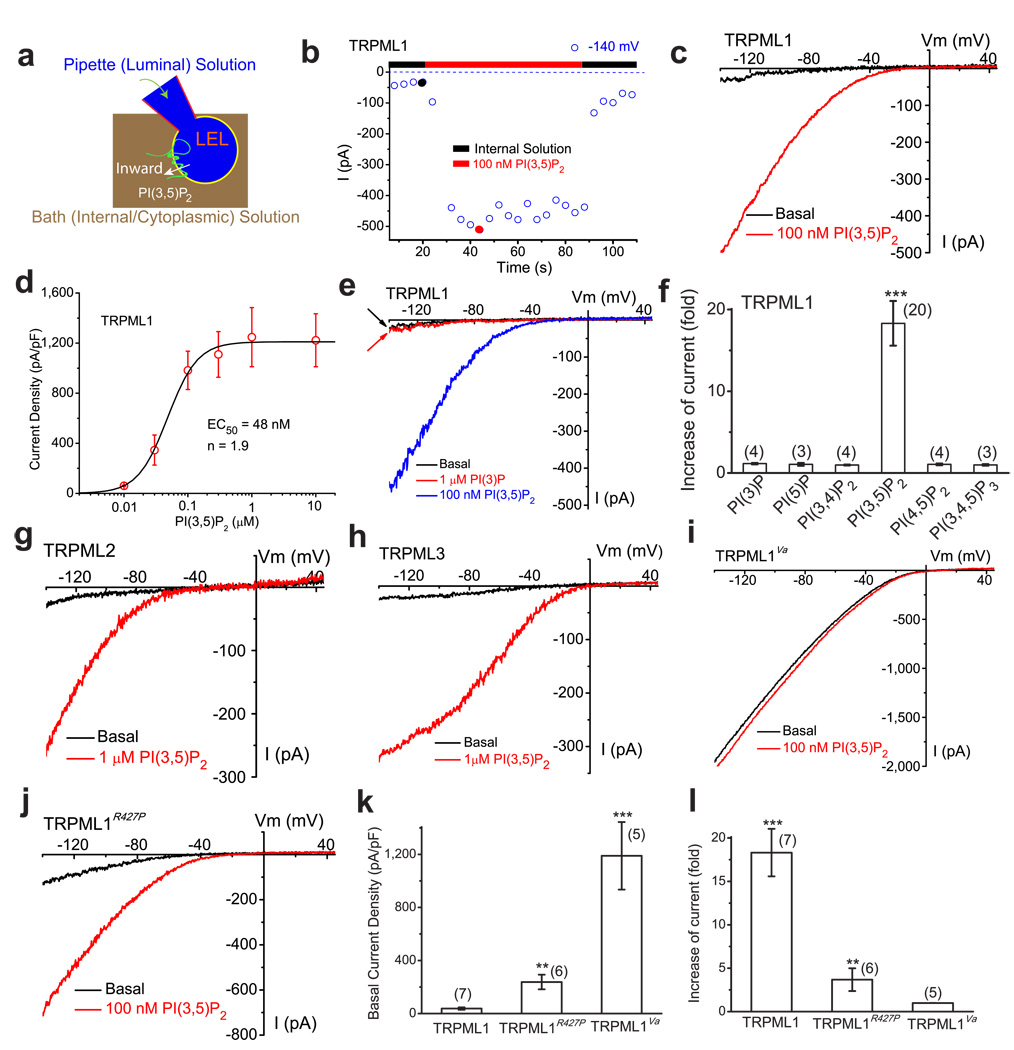
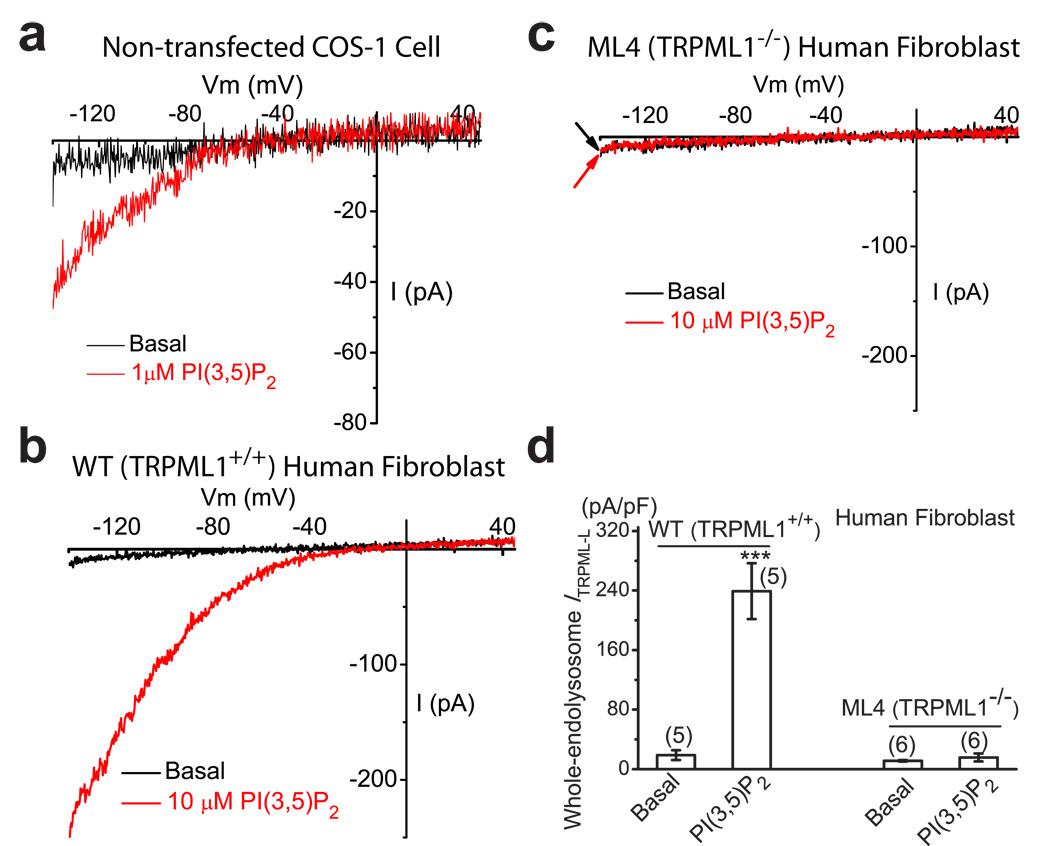
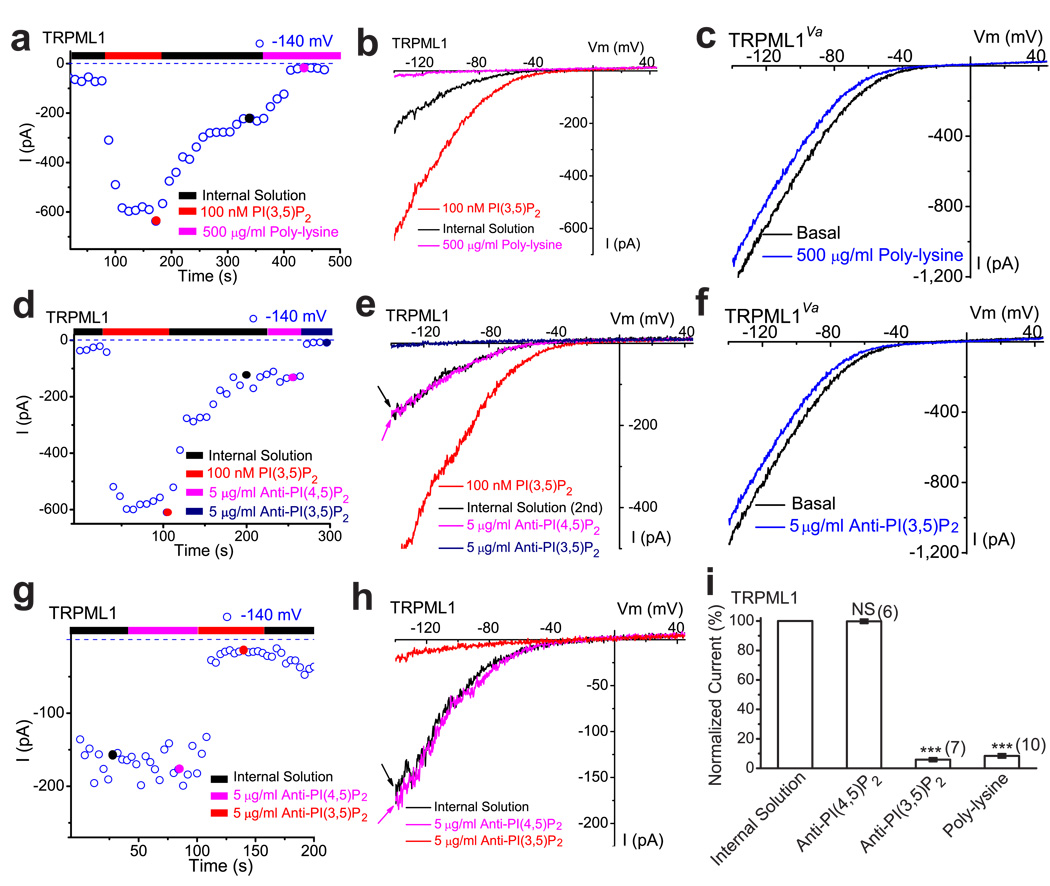
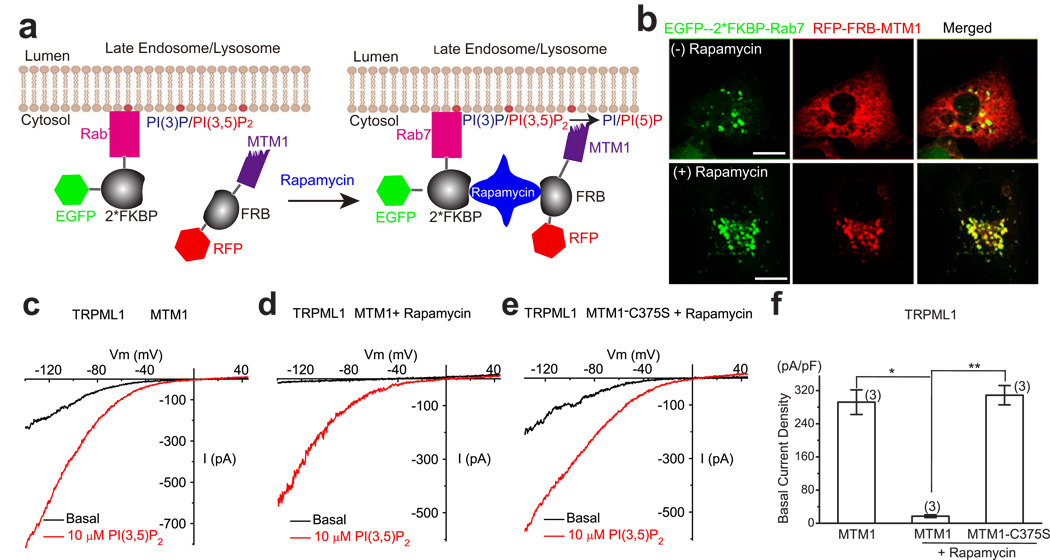
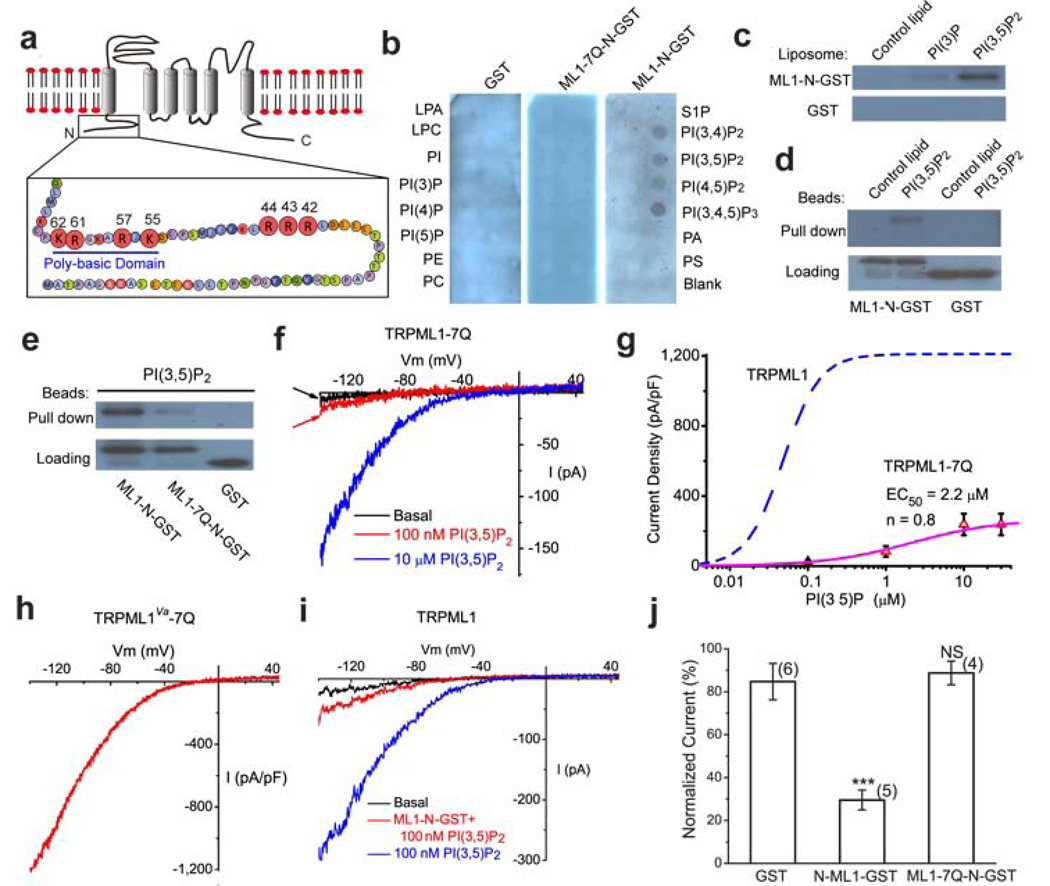
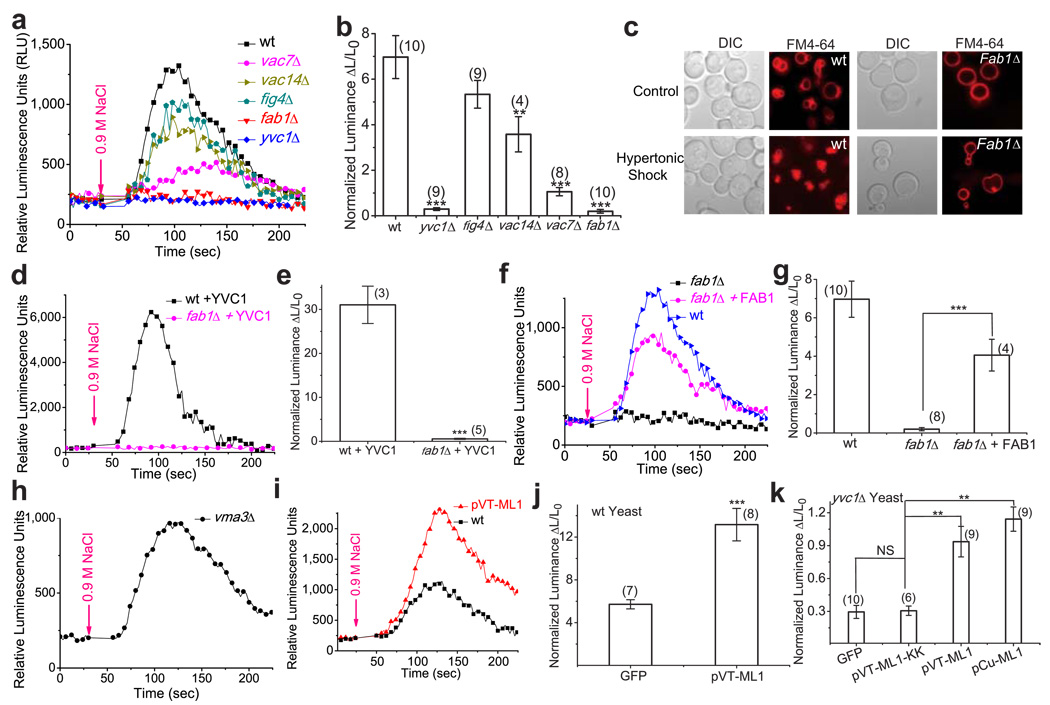
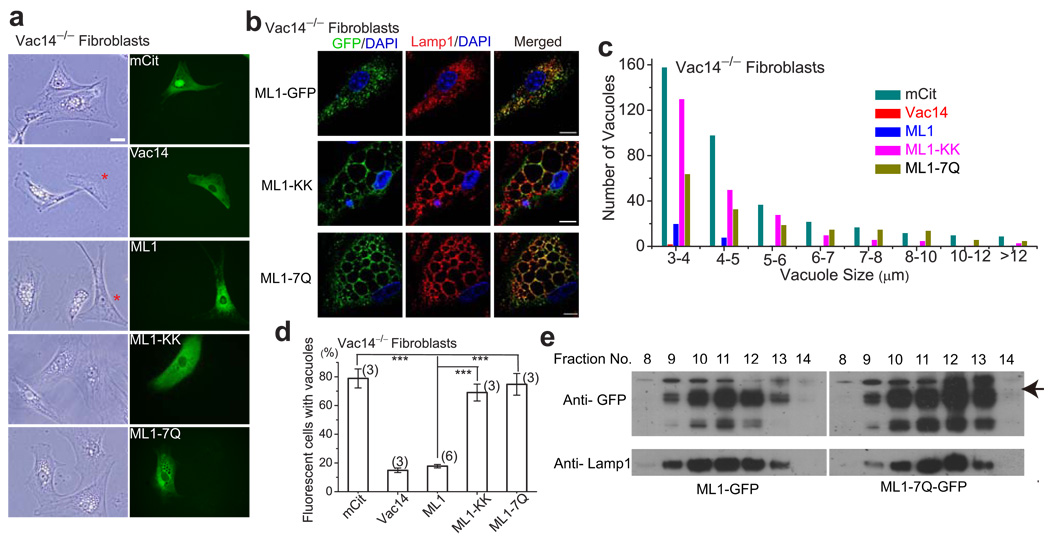
Membrane fusion and fission events in intracellular trafficking are controlled by both intraluminal Ca(2+) release and phosphoinositide (PIP) signalling. However, the molecular identities of the Ca(2+) release channels and the target proteins of PIPs are elusive. In this paper, by direct patch-clamping of the endolysosomal membrane, we report that PI(3,5)P(2), an endolysosome-specific PIP, binds and activates endolysosome-localized mucolipin transient receptor potential (TRPML) channels with specificity and potency. Both PI(3,5)P(2)-deficient cells and cells that lack TRPML1 exhibited enlarged endolysosomes/vacuoles and trafficking defects in the late endocytic pathway. We find that the enlarged vacuole phenotype observed in PI(3,5)P(2)-deficient mouse fibroblasts is suppressed by overexpression of TRPML1. Notably, this PI(3,5)P(2)-dependent regulation of TRPML1 is evolutionarily conserved. In budding yeast, hyperosmotic stress induces Ca(2+) release from the vacuole. In this study, we show that this release requires both PI(3,5)P(2) production and a yeast functional TRPML homologue. We propose that TRPMLs regulate membrane trafficking by transducing information regarding PI(3,5)P(2) levels into changes in juxtaorganellar Ca(2+), thereby triggering membrane fusion/fission events.
Keywords: Ca2+ release channel; Fab1; PI(3,5)P2; PIKfyve; TRP channel; Whole-endolysosome recording; endosome; lysosome; membrane trafficking; phosphoinositide; type IV Mucolipidosis; vacuole.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
