

Review
doi: 10.1186/gb-2010-11-8-216.
Epub 2010 Aug 26.
Dosage compensation and the global re-balancing of aneuploid genomes
Affiliations
- PMID: 20804581
- PMCID: PMC2945780
- DOI: 10.1186/gb-2010-11-8-216
Item in Clipboard
Review
Dosage compensation and the global re-balancing of aneuploid genomes
Genome Biol.
2010.
Abstract
Diploid genomes are exquisitely balanced systems of gene expression. The dosage-compensation systems that evolved along with monosomic sex chromosomes exemplify the intricacies of compensating for differences in gene copy number by transcriptional regulation.
Figures
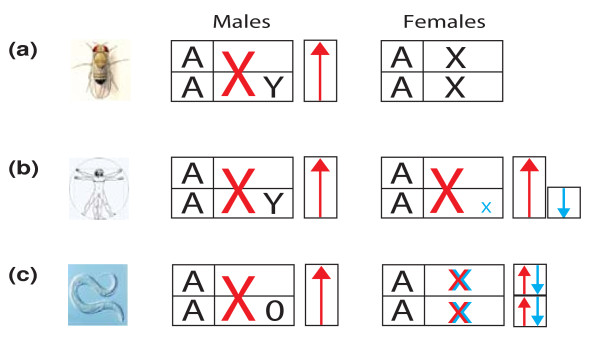
Schematic representation of different dosage-compensation systems. (a) Drosophila melanogaster, (b) Homo sapiens, (c) Caenorhabditis elegans. Combinations of chromosomes in the diploid somatic cells of males and females are shown. The sex chromosomes are symbolized by the letters X and Y, autosomes as A. Dosage-compensated chromosomes are colored: red indicates activation, blue repression. The sizes of the As indicate the average expression level of an autosome in a diploid cell. The sizes of the X chromosomes reflect their activity state (see text). The arrows represent the activating and repressive factors that determine the activity of the corresponding sex chromosome. In Drosophila (a), the male X chromosome is transcriptionally activated twofold in the male to match the total level of expression from the two female X chromosomes. In mammals (b), X chromosomes are hypertranscribed in both sexes, and to equalize X-chromosomal gene expression between the sexes, one of the two X chromosomes is inactivated in females. In C. elegans (c), males do not have a Y chromosome (O indicates its absence) and XX individuals are hermaphrodites. Worms also overexpress X-linked genes in a sex-independent manner, as indicated by the red-colored Xs, but subsequently halve the expression levels of the genes from both X chromosomes in the hermaphrodite (indicated by the blue Xs) to equalize gene dosage between the sexes.
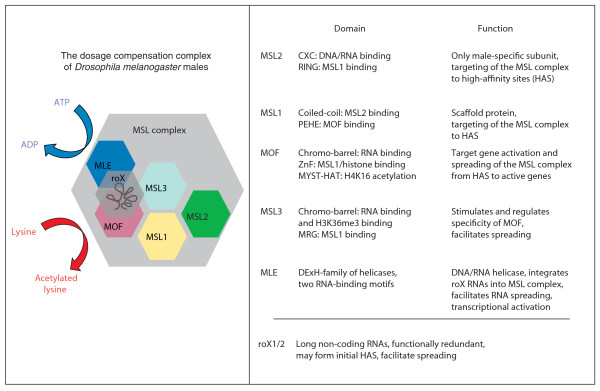
The Drosophila melanogaster male dosage-compensation complex. The complex, called the MSL complex in Drosophila, consists of five proteins (MSL1, MSL2, MSL3, MOF, MLE) and two non-coding roX RNAs. The proteins, but not the roX RNAs, are evolutionarily conserved, as related proteins can be found in yeast and humans (for details see [30,68,69]). The box lists the conserved protein domains of the individual members of the Drosophila MSL complex and their identified functions for dosage compensation. MSL2 is the only male-specific protein subunit; all other subunits are present in both sexes. The two roX RNAs (see bottom of table) are also only expressed in males. The curved arrows symbolize the known enzymatic activities in the dosage-compensation complex. MLE is an RNA helicase that hydrolyzes ATP to effect conformational changes in DNA and RNA [70]. MOF is a lysine acetyltransferase with specificity for lysine 16 of histone H4. Abbreviations of the protein domains are: CXC, cysteine-rich domain; ZnF, zinc finger; PEHE, proline-glutamic acid-histidine-glutamic acid; HAT, histone acetyltransferase; MYST, MOZ (monocytic leukemia zinc finger protein), YBF2/SAS3 (something about silencing 3), SAS2 and TIP60 (60 kDa Tat-interactive protein); MRG, mortality factor on chromosome 4 related gene and DExH, aspartic acid-glutamic acid-x-histidine.
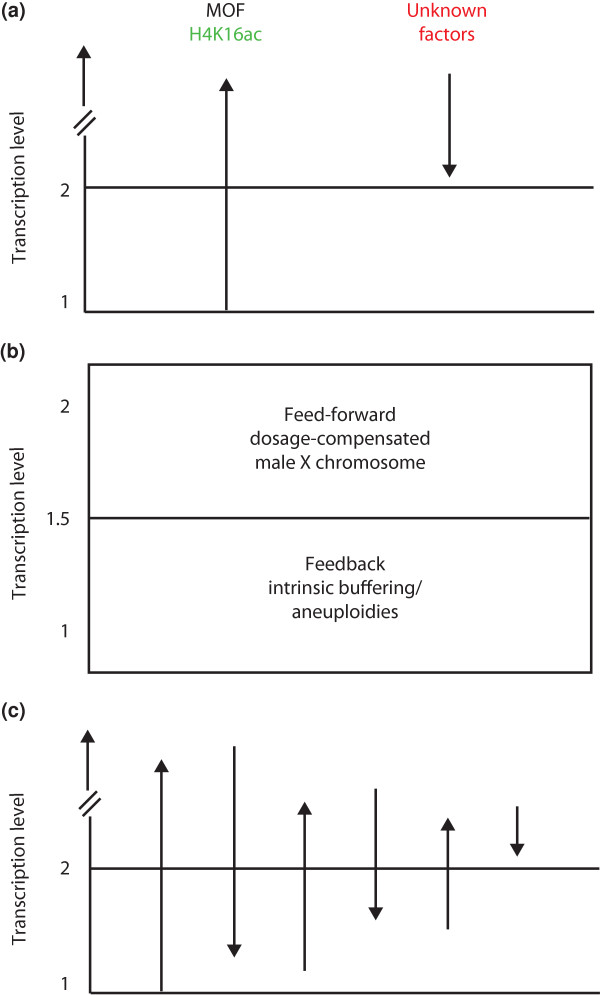
Possible mechanisms for dosage compensation. (a) The twofold activation of the single male X chromosome in Drosophila could be achieved by a large, MOF-dependent activation of transcription through H4K16 acetylation and its counteraction by yet unknown factors, mediated by the dosage-compensation complex in males [51]. In (a,b), transcriptional level 1 refers to the normal regulated level of transcription from a single uncompensated X chromosome in females. (b) Furthermore, the twofold activation of the male X chromosome could be achieved by a combination of mechanisms: a general buffering/feedback component and a dedicated feed-forward mechanism (dosage compensation as suggested in (a)) [7]. The effects of these two processes could add up to the expected twofold compensation required to equalize the expression of X-linked genes between the sexes. (c) Precise transcription levels could result from negotiation between a number of activating and repressive factors (up and down arrows). In this instance, transcriptional level 1 refers to a 'basal' transcription state. This hypothetical model assumes that additional factors beyond those mentioned in (a) and (b) contribute to final transcription levels, such as male-enriched protein kinases, heterochromatin components, chromatin remodelers, and others (for details, see text).
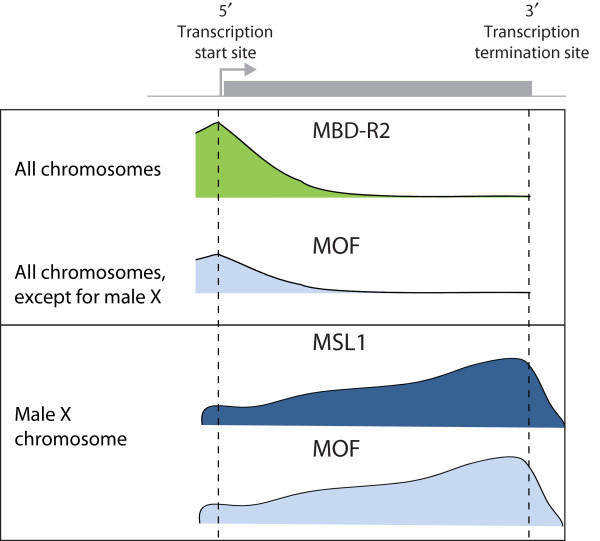
Schematic representation of the distribution of the key regulators of dosage compensation on a target gene in Drosophila. The gene is depicted as a gray bar at the top of the figure, with the arrow representing the transcription start site. The figure is based on genome-wide binding studies of MOF, MBD-R2 and MSL1. The upper panel shows that MBD-R2 is enriched at promoters (5') on all chromosomes in both sexes, underscoring its function as a general transcriptional facilitator. MOF co-localizes with the promoter peak of MBD-R2 on all chromosomes except for the male X chromosome, where it is more enriched towards the 3' end of the target gene as a result of its association with the dosage-compensation complex (bottom panel). The MSL1 profile serves as a marker for the presence of the dosage-compensation complex [51]. For details see text.
Similar articles
-
Dosage compensation of the sex chromosomes and autosomes.Semin Cell Dev Biol. 2016 Aug;56:9-18. doi: 10.1016/j.semcdb.2016.04.013. Epub 2016 Apr 22. Semin Cell Dev Biol. 2016. PMID: 27112542 Free PMC article. Review.
-
Dosage-dependent gene regulation in multicellular eukaryotes: implications for dosage compensation, aneuploid syndromes, and quantitative traits.Dev Biol. 2001 Jun 15;234(2):275-88. doi: 10.1006/dbio.2001.0262. Dev Biol. 2001. PMID: 11396999 Review.
-
Gene dosage compensation: Origins, criteria to identify compensated genes, and mechanisms including sensor loops as an emerging systems-level property in cancer.Cancer Med. 2023 Dec;12(24):22130-22155. doi: 10.1002/cam4.6719. Epub 2023 Nov 21. Cancer Med. 2023. PMID: 37987212 Free PMC article. Review.
-
Extensive protein dosage compensation in aneuploid human cancers.Genome Res. 2022 Jul;32(7):1254-1270. doi: 10.1101/gr.276378.121. Epub 2022 Jun 14. Genome Res. 2022. PMID: 35701073 Free PMC article.
-
Aneuploidy and gene expression: is there dosage compensation?Epigenomics. 2019 Dec;11(16):1827-1837. doi: 10.2217/epi-2019-0135. Epub 2019 Nov 22. Epigenomics. 2019. PMID: 31755744 Free PMC article. Review.
Cited by
-
Genome-wide haploinsufficiency screen reveals a novel role for γ-TuSC in spindle organization and genome stability.Mol Biol Cell. 2013 Sep;24(17):2753-63. doi: 10.1091/mbc.E12-12-0902. Epub 2013 Jul 3. Mol Biol Cell. 2013. PMID: 23825022 Free PMC article.
-
New insights into the troubles of aneuploidy.Annu Rev Cell Dev Biol. 2012;28:189-214. doi: 10.1146/annurev-cellbio-101011-155807. Epub 2012 Jul 9. Annu Rev Cell Dev Biol. 2012. PMID: 22804579 Free PMC article. Review.
-
Buffering and the evolution of chromosome-wide gene regulation.Chromosoma. 2011 Jun;120(3):213-25. doi: 10.1007/s00412-011-0319-8. Epub 2011 Apr 20. Chromosoma. 2011. PMID: 21505791 Free PMC article. Review.
-
Transcriptional Aneuploidy Responses of Brassica rapa-oleracea Monosomic Alien Addition Lines (MAALs) Derived From Natural Allopolyploid B. napus.Front Genet. 2019 Feb 13;10:67. doi: 10.3389/fgene.2019.00067. eCollection 2019. Front Genet. 2019. PMID: 30815011 Free PMC article.
-
X chromosome regulation: diverse patterns in development, tissues and disease.Nat Rev Genet. 2014 Jun;15(6):367-78. doi: 10.1038/nrg3687. Epub 2014 Apr 15. Nat Rev Genet. 2014. PMID: 24733023 Free PMC article. Review.
References
-
- Schinzel A. Catalogue of Unbalanced Chromosome Aberrations in Man. Berlin: Walter de Gruyter; 2001.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
