

Polycomb group proteins: multi-faceted regulators of somatic stem cells and cancer
- PMID: 20804967
- PMCID: PMC4959883
- DOI: 10.1016/j.stem.2010.08.002
Polycomb group proteins: multi-faceted regulators of somatic stem cells and cancer
Abstract
Polycomb Group (PcG) proteins are transcriptional repressors that epigenetically modify chromatin and participate in the establishment and maintenance of cell fates. These proteins play important roles in both stem cell self-renewal and in cancer development. Our understanding of their mechanism of action has greatly advanced over the past 10 years, but many unanswered questions remain. In this review, we present the currently available experimental data that connect PcG protein function with some of the key processes which govern somatic stem cell activity. We also highlight recent studies suggesting that a delicate balance in PcG gene dosage is crucial for proper stem cell homeostasis and prevention of cancer stem cell development.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
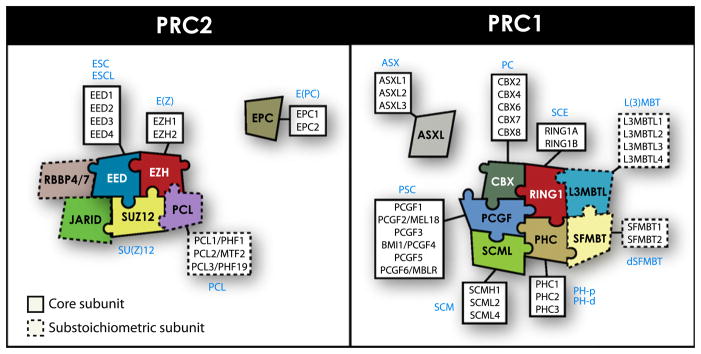

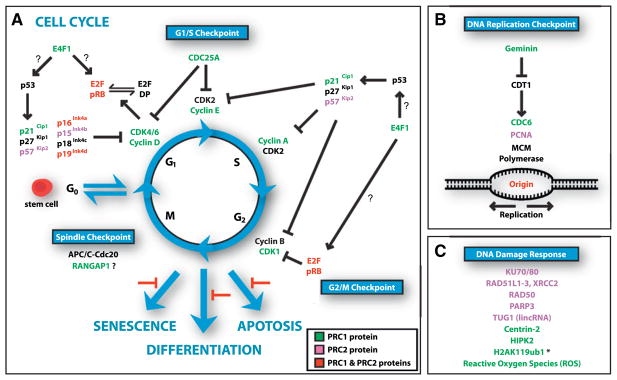
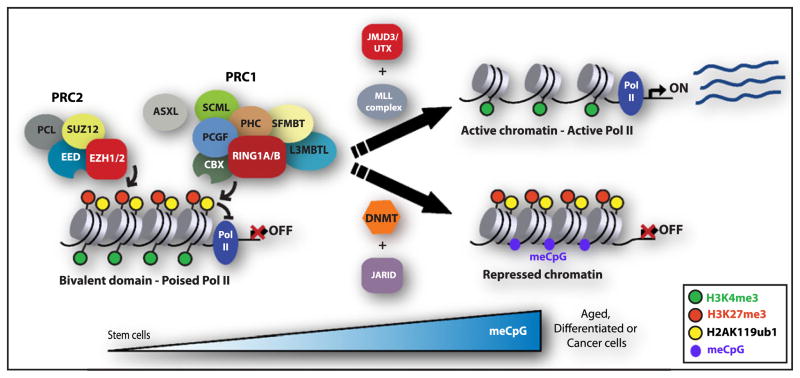
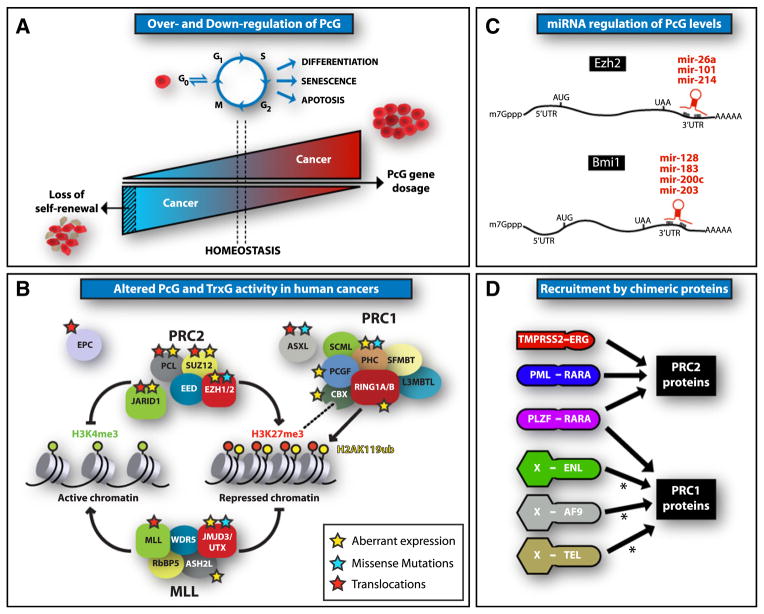
References
-
- Akasaka T, Takahashi N, Suzuki M, Koseki H, Bodmer R, Koga H. MBLR, a new RING finger protein resembling mammalian Polycomb gene products, is regulated by cell cycle-dependent phosphorylation. Genes Cells. 2002;7:835–850. - PubMed
-
- Attwooll C, Oddi S, Cartwright P, Prosperini E, Agger K, Steensgaard P, Wagener C, Sardet C, Moroni MC, Helin K. A novel repressive E2F6 complex containing the polycomb group protein, EPC1, that interacts with EZH2 in a proliferation-specific manner. J Biol Chem. 2005;280:1199–1208. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
