

Stat3 activation is limiting for reprogramming to ground state pluripotency
- PMID: 20804969
- PMCID: PMC3459098
- DOI: 10.1016/j.stem.2010.06.022
Stat3 activation is limiting for reprogramming to ground state pluripotency
Abstract
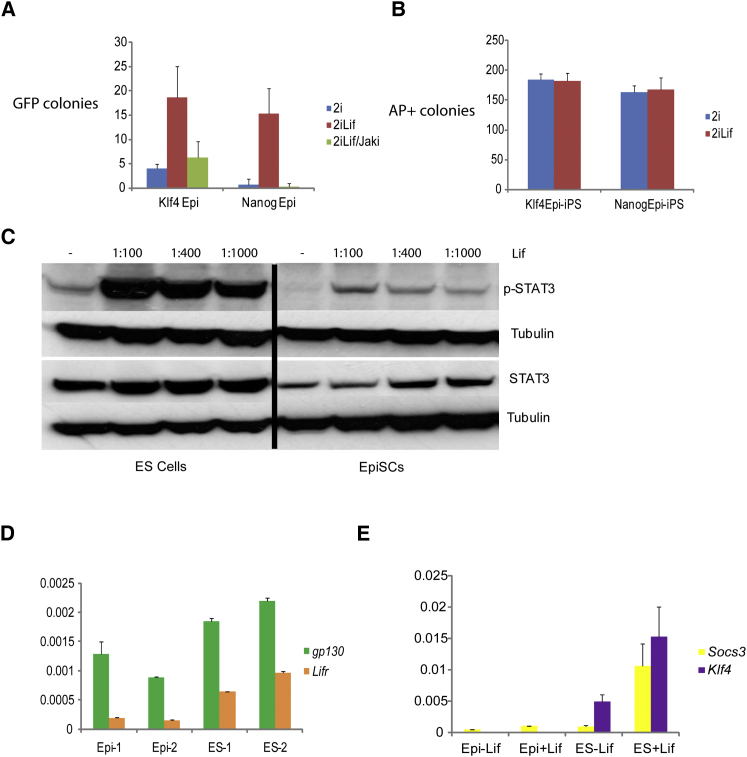
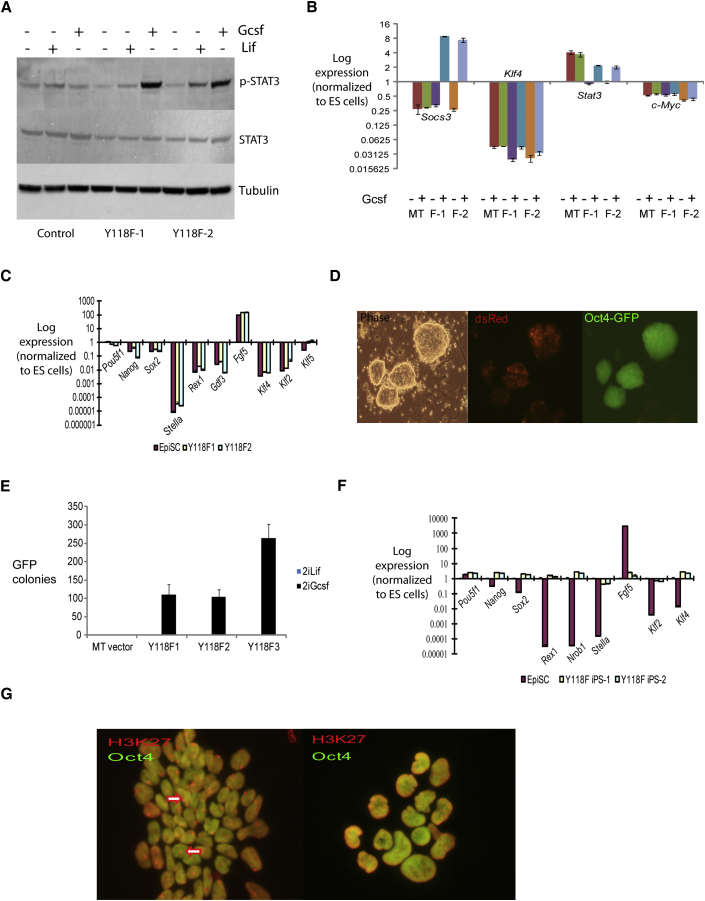
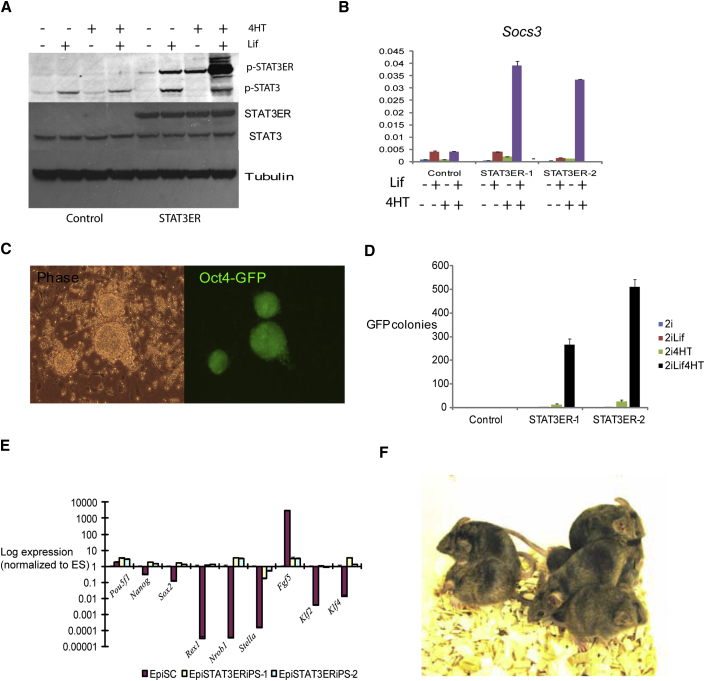
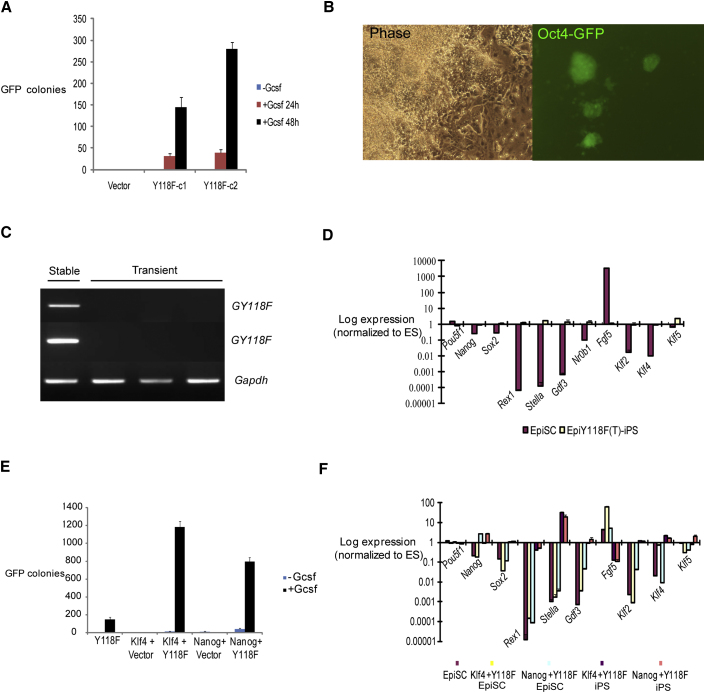
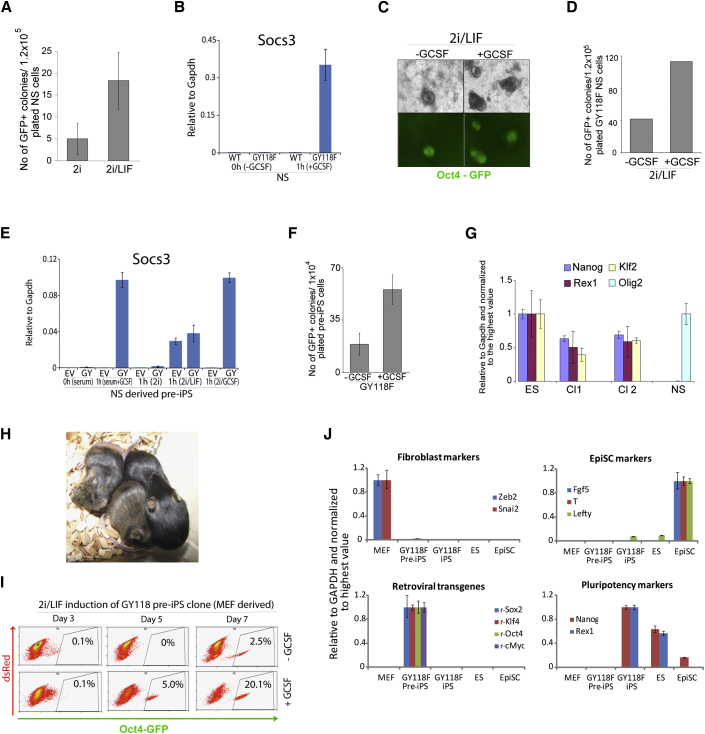
The cytokine leukemia inhibitory factor (Lif) sustains self-renewal of mouse embryonic and induced pluripotent stem cells by activating Jak kinase and the transcription factor Stat3. Here we investigate whether Jak/Stat3 may also contribute to induction of pluripotency. EpiSCs derived from postimplantation embryos express low levels of Lif receptor and Stat3. We introduced into EpiSCs a Jak/Stat3 activating receptor (GY118F) responsive to granulocyte colony stimulating factor (Gcsf). On transfer to ground state culture, in which MAPK signaling and glycogen synthase kinase are inhibited, Gcsf induced transcriptional resetting and functional reprogramming. Activation of a tamoxifen-regulatable fusion, Stat3ER(T2), also converted EpiSCs into chimera-competent iPSCs. We exploited GY118F to increase Jak/Stat3 activity during somatic cell reprogramming. Incompletely reprogrammed cells derived from neural stem cells or fibroblasts responded to Gcsf with elevated frequencies of progression to ground state pluripotency. These findings indicate that Jak/Stat3 participate directly in molecular reprogramming and that activation of this pathway is a limiting component.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
Comment in
-
The STATs on naive iPSC reprogramming.Cell Stem Cell. 2010 Sep 3;7(3):274-6. doi: 10.1016/j.stem.2010.08.005. Cell Stem Cell. 2010. PMID: 20804962 No abstract available.
References
-
- Batlle-Morera L., Smith A., Nichols J. Parameters influencing derivation of embryonic stem cells from murine embryos. Genesis. 2008;46:758–767. - PubMed
-
- Bourillot P.Y., Aksoy I., Schreiber V., Wianny F., Schulz H., Hummel O., Hubner N., Savatier P. Novel STAT3 target genes exert distinct roles in the inhibition of mesoderm and endoderm differentiation in cooperation with Nanog. Stem Cells. 2009;27:1760–1771. - PubMed
-
- Bradley A., Evans M.J., Kaufman M.H., Robertson E. Formation of germ-line chimaeras from embryo-derived teratocarcinoma cell lines. Nature. 1984;309:255–256. - PubMed
-
- Brons I.G., Smithers L.E., Trotter M.W., Rugg-Gunn P., Sun B., Chuva de Sousa Lopes S.M., Howlett S.K., Clarkson A., Ahrlund-Richter L., Pedersen R.A. Derivation of pluripotent epiblast stem cells from mammalian embryos. Nature. 2007;448:191–195. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
