

The distinct metabolic profile of hematopoietic stem cells reflects their location in a hypoxic niche
- PMID: 20804973
- PMCID: PMC4159713
- DOI: 10.1016/j.stem.2010.07.011
The distinct metabolic profile of hematopoietic stem cells reflects their location in a hypoxic niche
Abstract
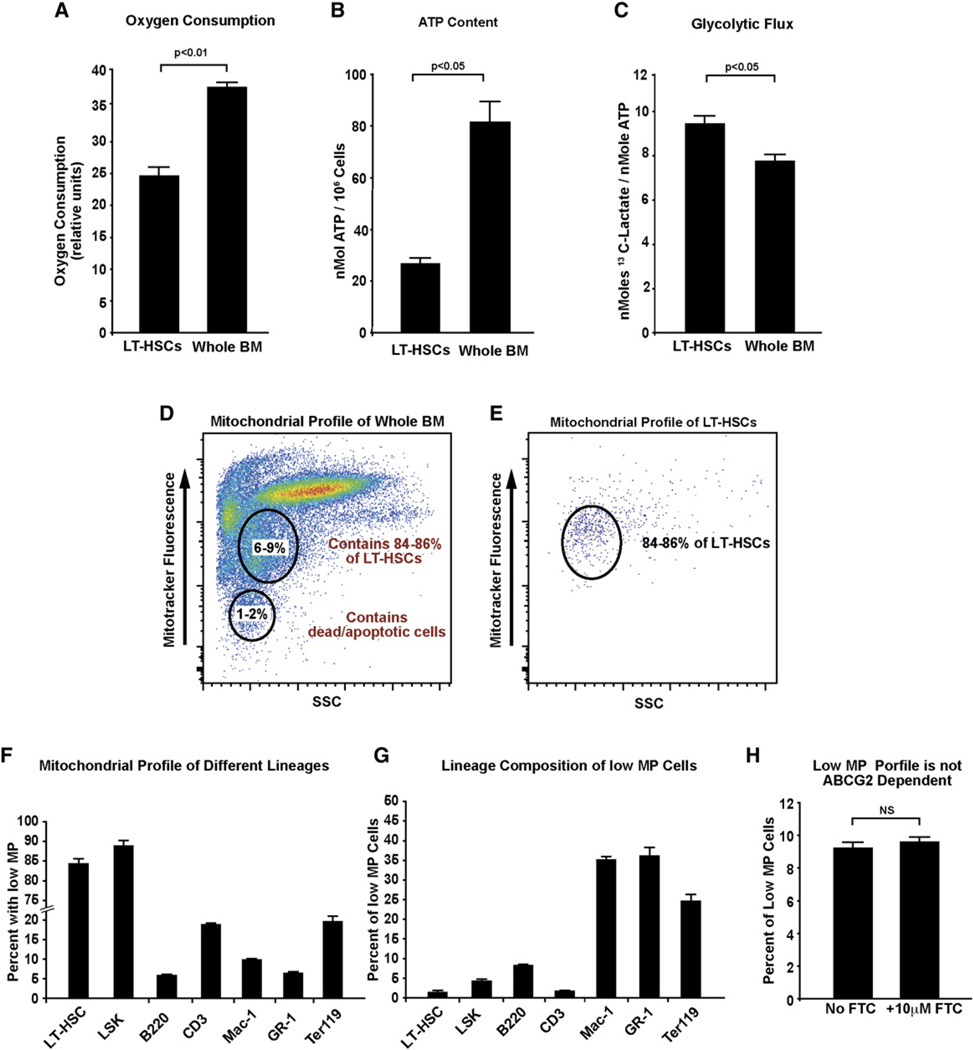
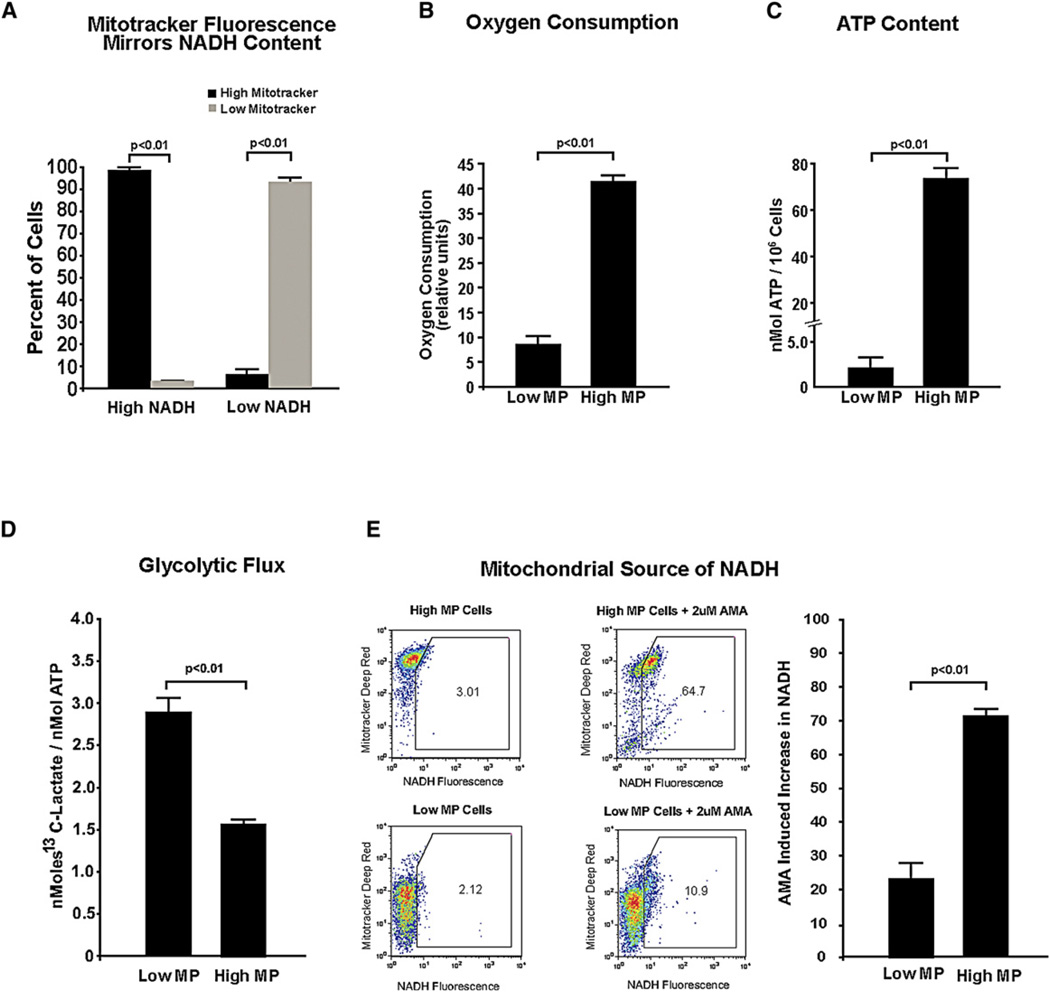
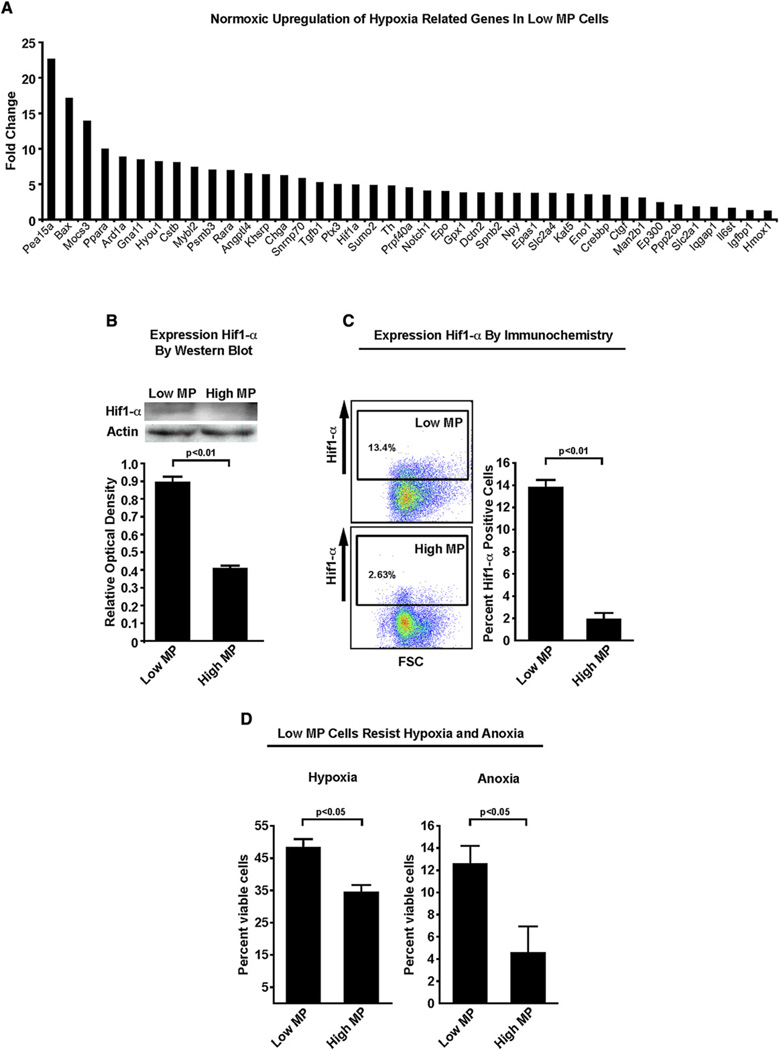
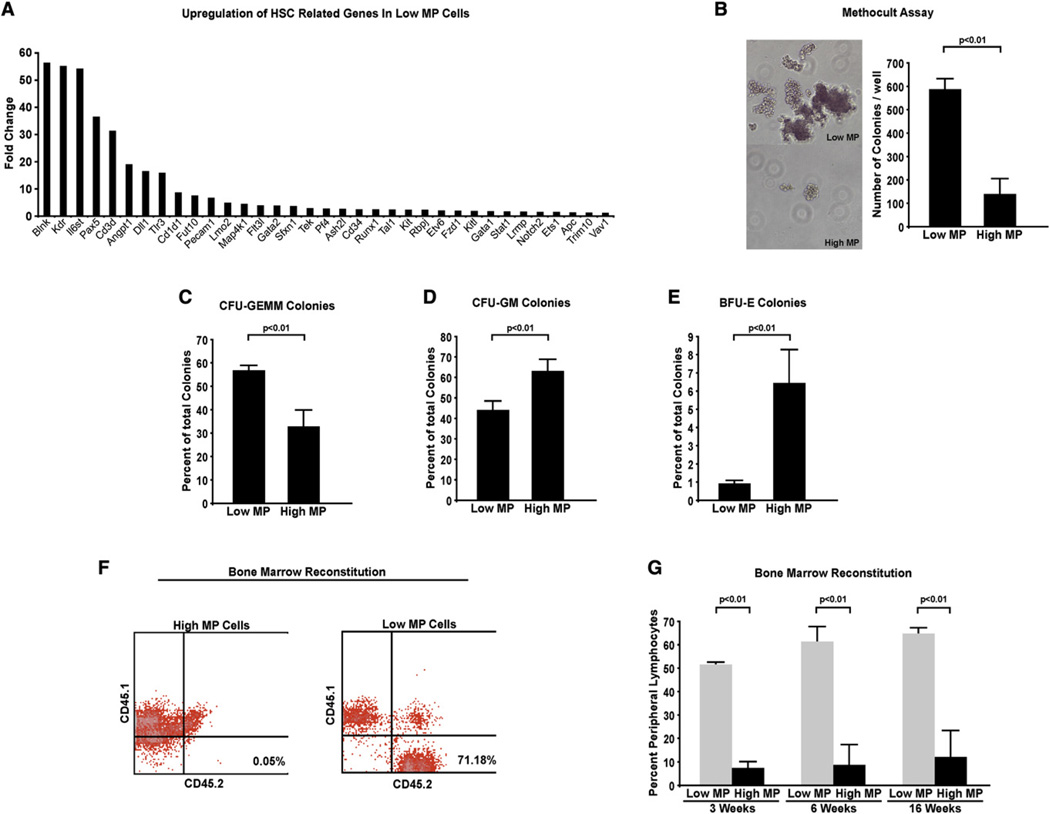
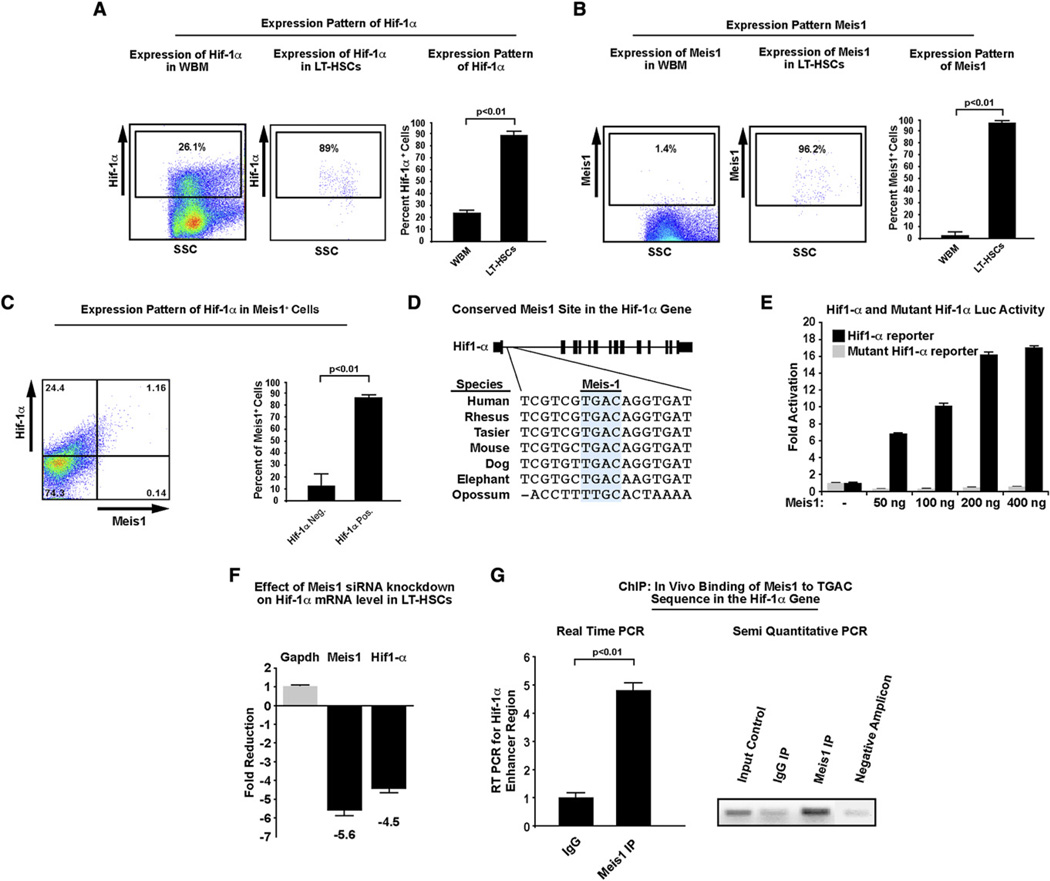
Bone marrow transplantation is the primary therapy for numerous hematopoietic disorders. The efficiency of bone marrow transplantation depends on the function of long-term hematopoietic stem cells (LT-HSCs), which is markedly influenced by their hypoxic niche. Survival in this low-oxygen microenvironment requires significant metabolic adaptation. Here, we show that LT-HSCs utilize glycolysis instead of mitochondrial oxidative phosphorylation to meet their energy demands. We used flow cytometry to identify a unique low mitochondrial activity/glycolysis-dependent subpopulation that houses the majority of hematopoietic progenitors and LT-HSCs. Finally, we demonstrate that Meis1 and Hif-1alpha are markedly enriched in LT-HSCs and that Meis1 regulates HSC metabolism through transcriptional activation of Hif-1alpha. These findings reveal an important transcriptional network that regulates HSC metabolism.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Hypoxia signaling in hematopoietic stem cells: a double-edged sword.Cell Stem Cell. 2010 Sep 3;7(3):276-8. doi: 10.1016/j.stem.2010.08.006. Cell Stem Cell. 2010. PMID: 20804963 No abstract available.
References
-
- Argiropoulos B, Yung E, Humphries RK. Unraveling the crucial roles of Meis1 in leukemogenesis and normal hematopoiesis. Genes Dev. 2007;21:2845–2849. - PubMed
-
- Azcoitia V, Aracil M, Martínez-A C, Torres M. The homeodomain protein Meis1 is essential for definitive hematopoiesis and vascular patterning in the mouse embryo. Dev. Biol. 2005;280:307–320. - PubMed
-
- Belaiba RS, Bonello S, Zähringer C, Schmidt S, Hess J, Kietzmann T, Görlach A. Hypoxia up-regulates hypoxia-inducible factor-1alpha transcription by involving phosphatidylinositol 3-kinase and nuclear factor kappaB in pulmonary artery smooth muscle cells. Mol. Biol. Cell. 2007;18:4691–4697. - PMC - PubMed
-
- Blouin CC, Pagé EL, Soucy GM, Richard DE. Hypoxic gene activation by lipopolysaccharide in macrophages: Implication of hypoxia-inducible factor 1alpha. Blood. 2004;103:1124–1130. - PubMed
-
- Bradley TR, Hodgson GS, Rosendaal M. The effect of oxygen tension on haemopoietic and fibroblast cell proliferation in vitro. J. Cell. Physiol. 1978;97(3 Pt 2) Suppl 1:517–522. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
