

Superoxide differentially controls pulmonary and systemic vascular tone through multiple signalling pathways
- PMID: 20805095
- PMCID: PMC3002873
- DOI: 10.1093/cvr/cvq275
Superoxide differentially controls pulmonary and systemic vascular tone through multiple signalling pathways
Abstract
Aims: the aim of this study was to determine the relative importance of Ca(2+) sensitization, ion channels, and intracellular Ca(2+) ([Ca(2+)](i)) in the mixed constrictor/relaxation actions of superoxide anion on systemic and pulmonary arteries.
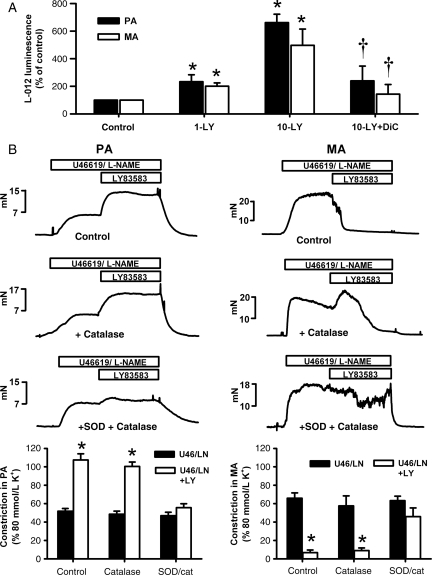
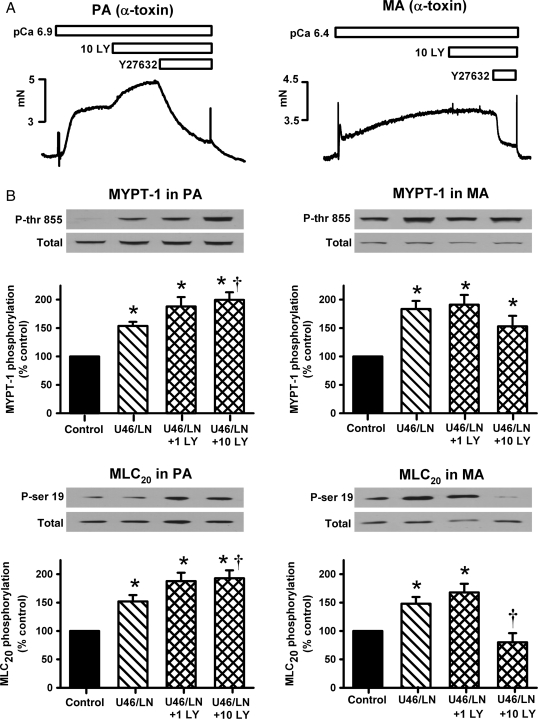
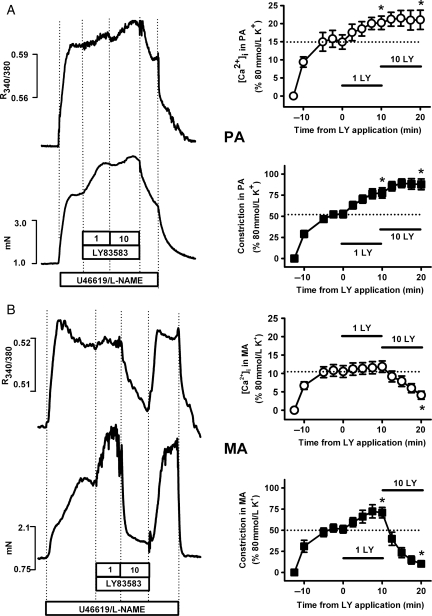
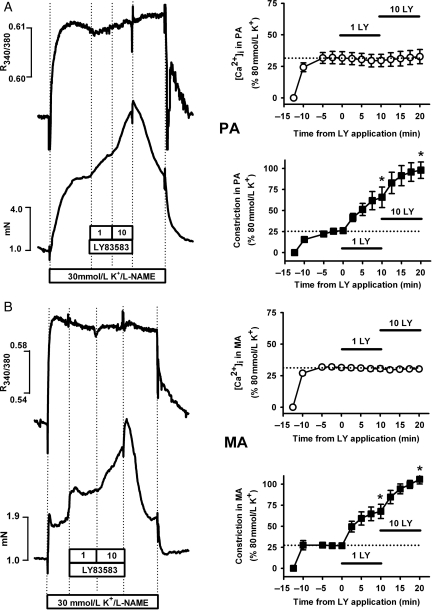
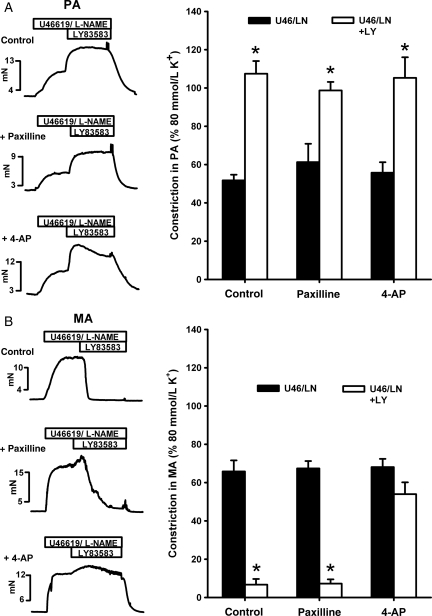
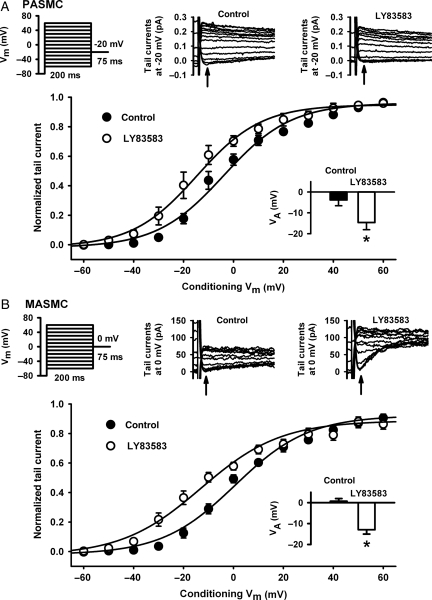
Methods and results: pulmonary and mesenteric arteries were obtained from rat. Superoxide was generated in arteries and cells with 6-anilino-5,8-quinolinequinone (LY83583). Following pre-constriction with U46619, 10 μmol/L LY83583 caused constriction in pulmonary and relaxation in mesenteric arteries. Both constrictor and relaxant actions of LY83583 were inhibited by superoxide dismutase and catalase. LY83583 caused Rho-kinase-dependent constriction in α-toxin-permeabilized pulmonary but not mesenteric arteries. Phosphorylation of myosin phosphatase-targeting subunit-1 (MYPT-1; as determined by western blot), was enhanced by LY83583 in pulmonary artery only. However, in both artery types, changes in tension were closely correlated with changes in phosphorylation of the 20 kDa myosin light chain as well as changes in [Ca(2+)](i) (as measured with Fura PE-3), with LY83583 causing increases in pulmonary and decreases in mesenteric arteries. When U46619 was replaced by 30 mmol/L K(+), all changes in [Ca(2+)](i) were abolished and LY83583 constricted both artery types. The K(V) channel inhibitor 4-aminopyridine abolished the LY83583-induced relaxation in mesenteric artery without affecting constriction in pulmonary artery. However, LY83583 caused a similar hyperpolarizing shift in the steady-state activation of K(V) current in isolated smooth muscle cells of both artery types.
Conclusions: superoxide only causes Rho-kinase-dependent Ca(2+) sensitization in pulmonary artery, resulting in constriction, and whilst it opens K(V) channels in both artery types, this only results in relaxation in mesenteric.
Figures
Similar articles
-
Superoxide constricts rat pulmonary arteries via Rho-kinase-mediated Ca(2+) sensitization.Free Radic Biol Med. 2009 Mar 1;46(5):633-42. doi: 10.1016/j.freeradbiomed.2008.11.015. Epub 2008 Dec 6. Free Radic Biol Med. 2009. PMID: 19103285 Free PMC article.
-
ROS-dependent activation of RhoA/Rho-kinase in pulmonary artery: Role of Src-family kinases and ARHGEF1.Free Radic Biol Med. 2017 Sep;110:316-331. doi: 10.1016/j.freeradbiomed.2017.06.022. Epub 2017 Jul 1. Free Radic Biol Med. 2017. PMID: 28673614 Free PMC article.
-
Thromboxane A2-induced inhibition of voltage-gated K+ channels and pulmonary vasoconstriction: role of protein kinase Czeta.Circ Res. 2003 Oct 3;93(7):656-63. doi: 10.1161/01.RES.0000095245.97945.FE. Epub 2003 Sep 11. Circ Res. 2003. PMID: 12970114
-
Postnatal maturational shift from PKCzeta and voltage-gated K+ channels to RhoA/Rho kinase in pulmonary vasoconstriction.Cardiovasc Res. 2005 Apr 1;66(1):84-93. doi: 10.1016/j.cardiores.2004.12.019. Epub 2005 Jan 27. Cardiovasc Res. 2005. PMID: 15769451
-
[Pulmonary hypertension: calcium channel-mediated signaling, present and future pharmacological targets].Rev Med Chil. 2023 Jun;151(6):753-763. doi: 10.4067/s0034-98872023000600753. Rev Med Chil. 2023. PMID: 38801384 Review. Spanish.
Cited by
-
Sphingosylphosphorylcholine potentiates vasoreactivity and voltage-gated Ca2+ entry via NOX1 and reactive oxygen species.Cardiovasc Res. 2015 Apr 1;106(1):121-30. doi: 10.1093/cvr/cvv029. Epub 2015 Feb 6. Cardiovasc Res. 2015. PMID: 25661082 Free PMC article.
-
Aging-associated oxidative stress leads to decrease in IAS tone via RhoA/ROCK downregulation.Am J Physiol Gastrointest Liver Physiol. 2014 Jun 1;306(11):G983-91. doi: 10.1152/ajpgi.00087.2014. Epub 2014 Apr 17. Am J Physiol Gastrointest Liver Physiol. 2014. PMID: 24742984 Free PMC article.
-
Perinatal hypoxia augments contractile impact of NADPH oxidase-derived ROS in early postnatal rat arteries.Pediatr Res. 2025 Feb;97(3):1220-1226. doi: 10.1038/s41390-024-03466-z. Epub 2024 Aug 10. Pediatr Res. 2025. PMID: 39127838
-
Hypoxic pulmonary vasoconstriction: mechanisms of oxygen-sensing.Curr Opin Anaesthesiol. 2011 Feb;24(1):13-20. doi: 10.1097/ACO.0b013e3283421201. Curr Opin Anaesthesiol. 2011. PMID: 21157304 Free PMC article. Review.
-
Bimodal effect of oxidative stress in internal anal sphincter smooth muscle.Am J Physiol Gastrointest Liver Physiol. 2015 Sep 1;309(5):G292-300. doi: 10.1152/ajpgi.00125.2015. Epub 2015 Jul 2. Am J Physiol Gastrointest Liver Physiol. 2015. PMID: 26138467 Free PMC article.
References
-
- Janssen-Heininger YM, Mossman BT, Heintz NH, Forman HJ, Kalyanaraman B, Finkel T, et al. Redox-based regulation of signal transduction: principles, pitfalls, and promises. Free Radic Biol Med. 2008;45:1–17. doi:10.1016/j.freeradbiomed.2008.03.011. - DOI - PMC - PubMed
-
- Shuvaev VV, Christofidou-Solomidou M, Bhora F, Laude K, Cai H, Dikalov S, et al. Targeted detoxification of selected reactive oxygen species in the vascular endothelium. J Pharmacol Exp Ther. 2009;331:404–411. doi:10.1124/jpet.109.156877. - DOI - PMC - PubMed
-
- Crosswhite P, Sun Z. Nitric oxide, oxidative stress and inflammation in pulmonary arterial hypertension. J Hypertens. 2010;28:201–212. doi:10.1097/HJH.0b013e328332bcdb. - DOI - PMC - PubMed
-
- Kamezaki F, Tasaki H, Yamashita K, Tsutsui M, Koide S, Nakata S, et al. Gene transfer of extracellular superoxide dismutase ameliorates pulmonary hypertension in rats. Am J Respir Crit Care Med. 2008;177:219–226. doi:10.1164/rccm.200702-264OC. - DOI - PubMed
-
- Gao YJ, Hirota S, Zhang DW, Janssen LJ, Lee RM. Mechanisms of hydrogen-peroxide-induced biphasic response in rat mesenteric artery. Br J Pharmacol. 2003;138:1085–1092. doi:10.1038/sj.bjp.0705147. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
