

The NuRD chromatin-remodeling complex regulates signaling and repair of DNA damage
- PMID: 20805320
- PMCID: PMC2935570
- DOI: 10.1083/jcb.201001048
The NuRD chromatin-remodeling complex regulates signaling and repair of DNA damage
Abstract
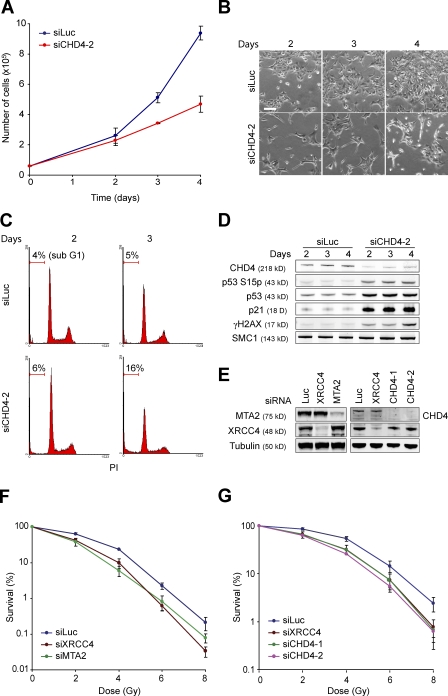
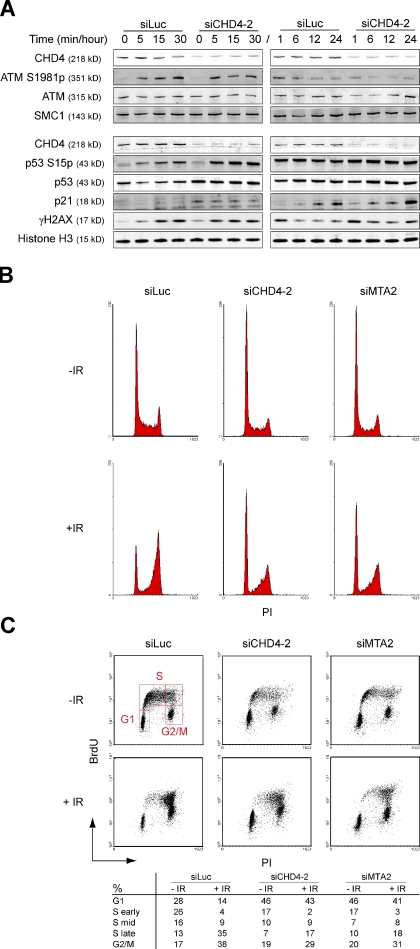
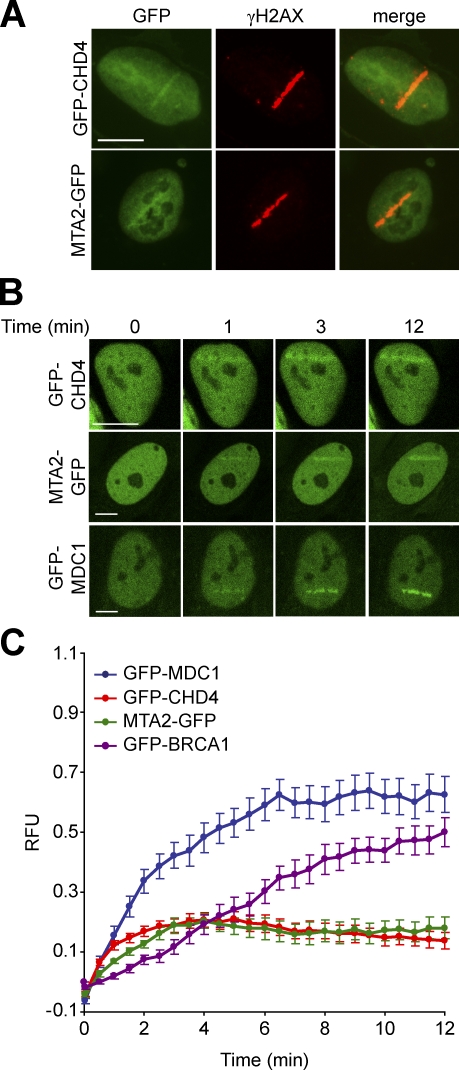
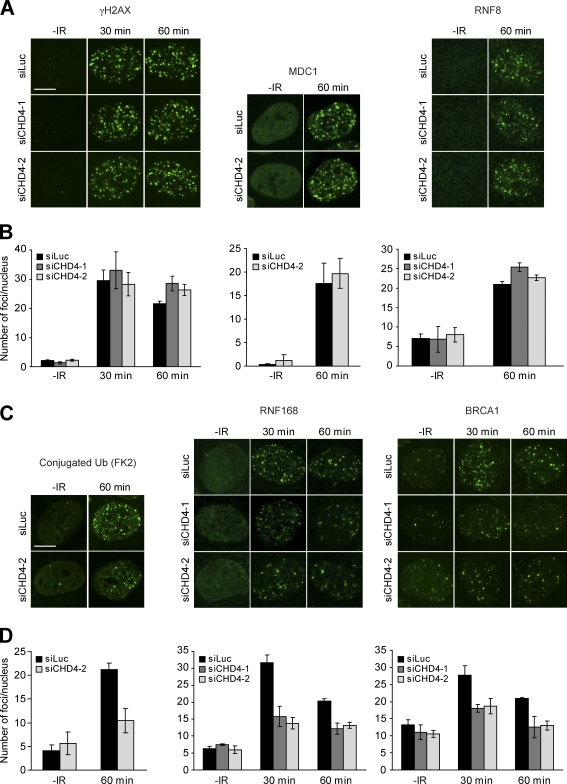
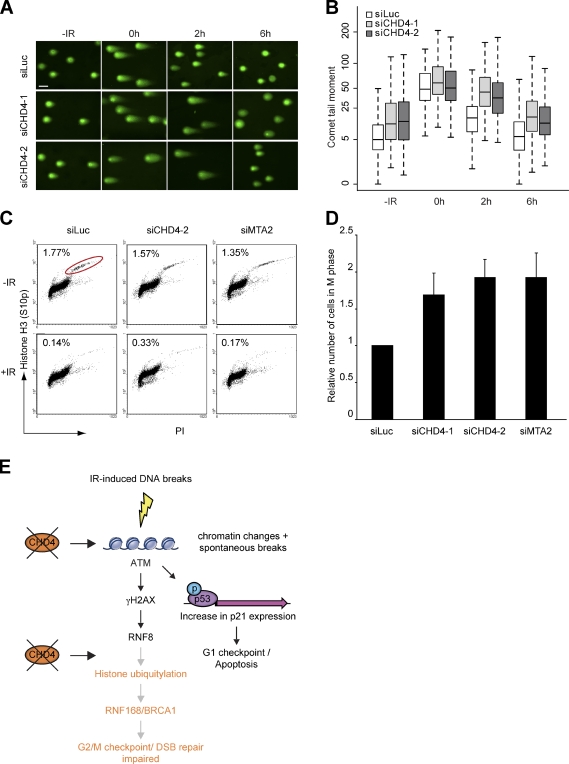
Cells respond to ionizing radiation (IR)-induced DNA double-strand breaks (DSBs) by orchestrating events that coordinate cell cycle progression and DNA repair. How cells signal and repair DSBs is not yet fully understood. A genome-wide RNA interference screen in Caenorhabditis elegans identified egr-1 as a factor that protects worm cells against IR. The human homologue of egr-1, MTA2 (metastasis-associated protein 2), is a subunit of the nucleosome-remodeling and histone deacetylation (NuRD) chromatin-remodeling complex. We show that knockdown of MTA2 and CHD4 (chromodomain helicase DNA-binding protein 4), the catalytic subunit (adenosine triphosphatase [ATPase]) of NuRD, leads to accumulation of spontaneous DNA damage and increased IR sensitivity. MTA2 and CHD4 accumulate in DSB-containing chromatin tracks generated by laser microirradiation. Directly at DSBs, CHD4 stimulates RNF8/RNF168-dependent formation of ubiquitin conjugates to facilitate the accrual of RNF168 and BRCA1. Finally, we show that CHD4 promotes DSB repair and checkpoint activation in response to IR. Thus, the NuRD chromatin-remodeling complex is a novel regulator of DNA damage responses that orchestrates proper signaling and repair of DSBs.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
