

Molecular insights into amyloid regulation by membrane cholesterol and sphingolipids: common mechanisms in neurodegenerative diseases
- PMID: 20807455
- PMCID: PMC2931503
- DOI: 10.1017/S1462399410001602
Molecular insights into amyloid regulation by membrane cholesterol and sphingolipids: common mechanisms in neurodegenerative diseases
Abstract
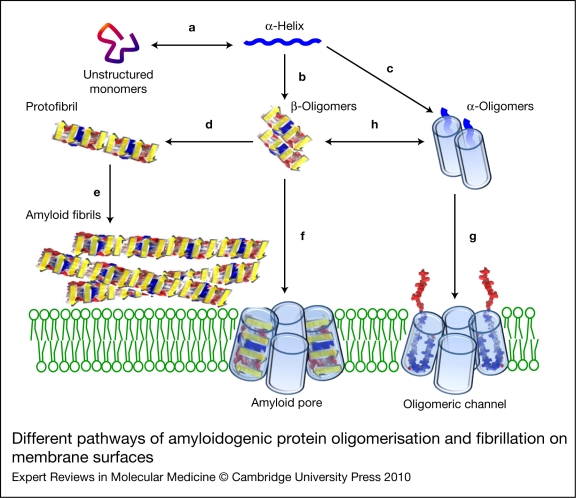
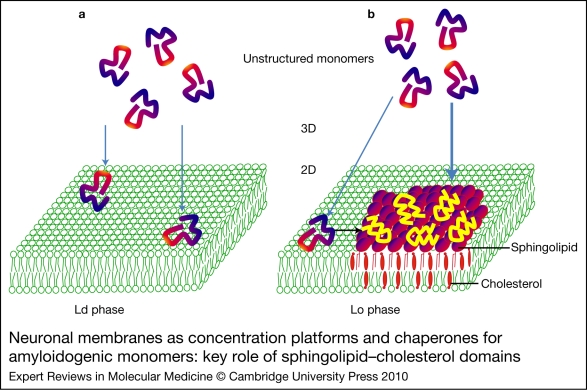
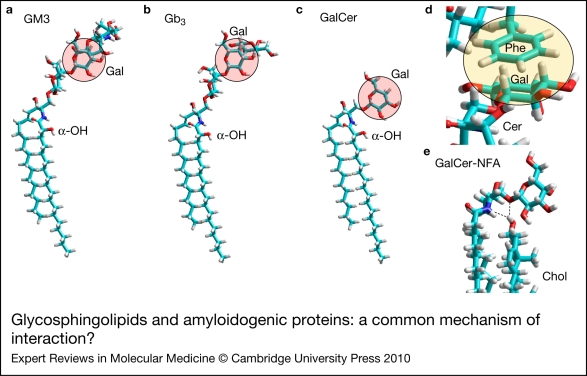
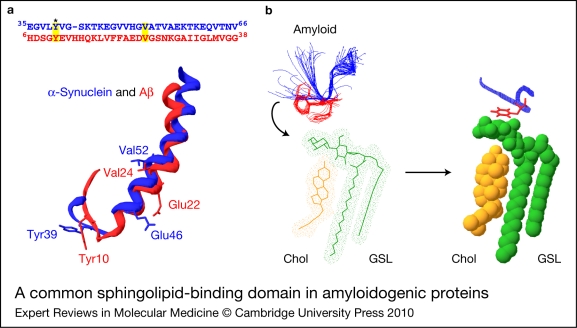
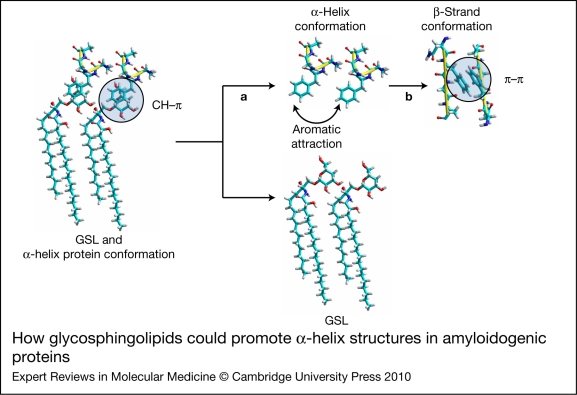
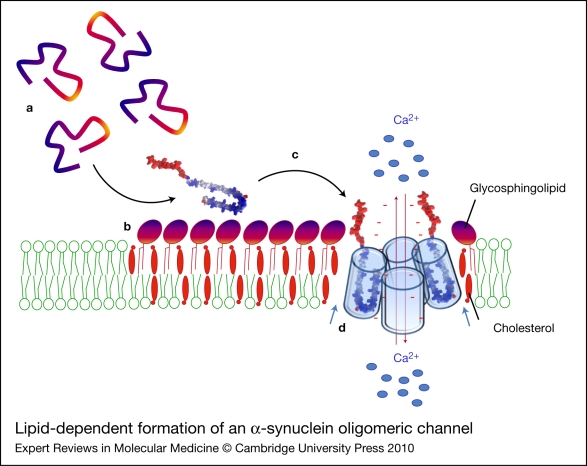
Alzheimer, Parkinson and other neurodegenerative diseases involve a series of brain proteins, referred to as 'amyloidogenic proteins', with exceptional conformational plasticity and a high propensity for self-aggregation. Although the mechanisms by which amyloidogenic proteins kill neural cells are not fully understood, a common feature is the concentration of unstructured amyloidogenic monomers on bidimensional membrane lattices. Membrane-bound monomers undergo a series of lipid-dependent conformational changes, leading to the formation of oligomers of varying toxicity rich in beta-sheet structures (annular pores, amyloid fibrils) or in alpha-helix structures (transmembrane channels). Condensed membrane nano- or microdomains formed by sphingolipids and cholesterol are privileged sites for the binding and oligomerisation of amyloidogenic proteins. By controlling the balance between unstructured monomers and alpha or beta conformers (the chaperone effect), sphingolipids can either inhibit or stimulate the oligomerisation of amyloidogenic proteins. Cholesterol has a dual role: regulation of protein-sphingolipid interactions through a fine tuning of sphingolipid conformation (indirect effect), and facilitation of pore (or channel) formation through direct binding to amyloidogenic proteins. Deciphering this complex network of molecular interactions in the context of age- and disease-related evolution of brain lipid expression will help understanding of how amyloidogenic proteins induce neural toxicity and will stimulate the development of innovative therapies for neurodegenerative diseases.
Figures
Similar articles
-
Deregulated sphingolipid metabolism and membrane organization in neurodegenerative disorders.Mol Neurobiol. 2010 Jun;41(2-3):314-40. doi: 10.1007/s12035-009-8096-6. Epub 2010 Feb 3. Mol Neurobiol. 2010. PMID: 20127207 Review.
-
Interaction of Alzheimer's β-amyloid peptides with cholesterol: mechanistic insights into amyloid pore formation.Biochemistry. 2014 Jul 22;53(28):4489-502. doi: 10.1021/bi500373k. Epub 2014 Jul 11. Biochemistry. 2014. PMID: 25000142 Review.
-
Elucidating the Structures of Amyloid Oligomers with Macrocyclic β-Hairpin Peptides: Insights into Alzheimer's Disease and Other Amyloid Diseases.Acc Chem Res. 2018 Mar 20;51(3):706-718. doi: 10.1021/acs.accounts.7b00554. Epub 2018 Mar 6. Acc Chem Res. 2018. PMID: 29508987 Free PMC article. Review.
-
Sphingolipids: membrane microdomains in brain development, function and neurological diseases.Open Biol. 2017 May;7(5):170069. doi: 10.1098/rsob.170069. Open Biol. 2017. PMID: 28566300 Free PMC article. Review.
-
Fluorescence as a method to reveal structures and membrane-interactions of amyloidogenic proteins.Biochim Biophys Acta. 2007 Aug;1768(8):1862-85. doi: 10.1016/j.bbamem.2007.03.015. Epub 2007 Mar 28. Biochim Biophys Acta. 2007. PMID: 17493579 Review.
Cited by
-
Cholesterol accelerates the binding of Alzheimer's β-amyloid peptide to ganglioside GM1 through a universal hydrogen-bond-dependent sterol tuning of glycolipid conformation.Front Physiol. 2013 Jun 10;4:120. doi: 10.3389/fphys.2013.00120. eCollection 2013. Front Physiol. 2013. PMID: 23772214 Free PMC article.
-
Challenges and approaches to understand cholesterol-binding impact on membrane protein function: an NMR view.Cell Mol Life Sci. 2018 Jun;75(12):2137-2151. doi: 10.1007/s00018-018-2789-9. Epub 2018 Mar 8. Cell Mol Life Sci. 2018. PMID: 29520423 Free PMC article. Review.
-
Structural dynamics of SARS-CoV-2 variants: A health monitoring strategy for anticipating Covid-19 outbreaks.J Infect. 2021 Aug;83(2):197-206. doi: 10.1016/j.jinf.2021.06.001. Epub 2021 Jun 3. J Infect. 2021. PMID: 34089757 Free PMC article.
-
Regulation of Functional Protein Aggregation by Multiple Factors: Implications for the Amyloidogenic Behavior of the CAP Superfamily Proteins.Int J Mol Sci. 2020 Sep 7;21(18):6530. doi: 10.3390/ijms21186530. Int J Mol Sci. 2020. PMID: 32906672 Free PMC article. Review.
-
Lipid rafts in neurodegeneration and neuroprotection.Mol Neurobiol. 2014 Aug;50(1):130-48. doi: 10.1007/s12035-013-8614-4. Epub 2013 Dec 22. Mol Neurobiol. 2014. PMID: 24362851 Review.
References
-
- Wright P.E., Dyson H.J.. Intrinsically unstructured proteins: re-assessing the protein structure-function paradigm. Journal of Molecular Biology. 1999;293:321–331. - PubMed
-
- Dunker A.K., Uversky V.N.. Signal transduction via unstructured protein conduits. Nature Chemical Biology. 2008;4:229–230. - PubMed
-
- Uversky V.N.. A protein-chameleon: conformational plasticity of alpha-synuclein, a disordered protein involved in neurodegenerative disorders. Journal of Biomolecular Structure and Dynamics. 2003;21:211–234. - PubMed
Further reading, resources and contacts
-
- Butterfield S.M., Lashuel H.A.. Amyloidogenic protein–membrane interactions: mechanistic insights from model systems. Angewandte Chemie 2010 . Jul 12; [Epub ahead of print]. - PubMed
-
This is a very complete and up-to-date review of the available data on the interaction of amyloidogenic proteins with model membranes.
-
- Matsuzaki K., Kato K., Yanagisawa K.. Abeta polymerization through interaction with membrane gangliosides. Biochimica et Biophysica Acta. 2010;1801:868–877. - PubMed
-
This excellent review explores the possibility that ganglioside binding is the initial and common step in the development of human diseases involving amyloidogenic proteins, including Alzheimer disease.
Websites
For a discussion on glycosphingolipid structure and conformation, see our GalCer website:
http://www.galcer.u-3mrs.fr/ The Alzheimer Research Forum is a very informative free website dedicated to understanding Alzheimer disease and related disorders, with resources on amyloidogenic proteins and mutations, and research news:
http://www.alzforum.org/ Useful resources on amyloid pores can be found at the following web page, held by Peter T. Lansbury, Jr:
http://lansbury.bwh.harvard.edu/amyloid_pore.htm
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
