

Crossveinless-2 is required for the relocalization of Chordin protein within the vertebral field in mouse embryos
- PMID: 20807528
- PMCID: PMC2975673
- DOI: 10.1016/j.ydbio.2010.08.025
Crossveinless-2 is required for the relocalization of Chordin protein within the vertebral field in mouse embryos
Abstract
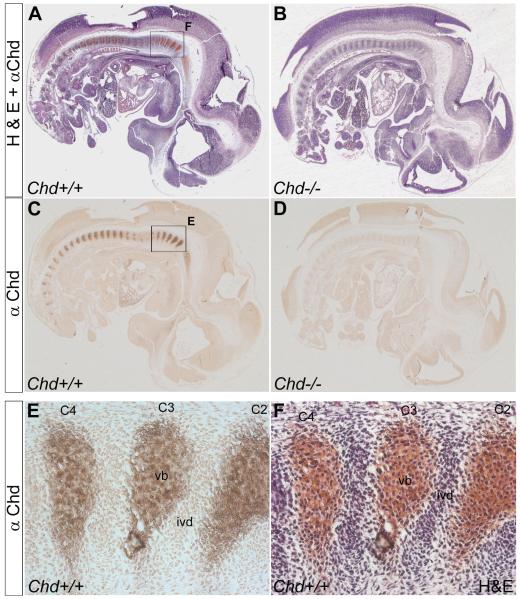
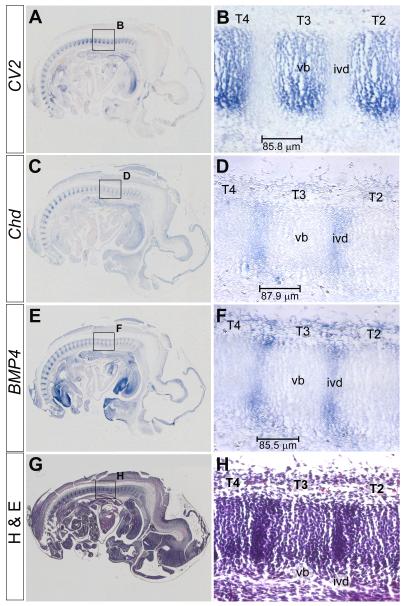
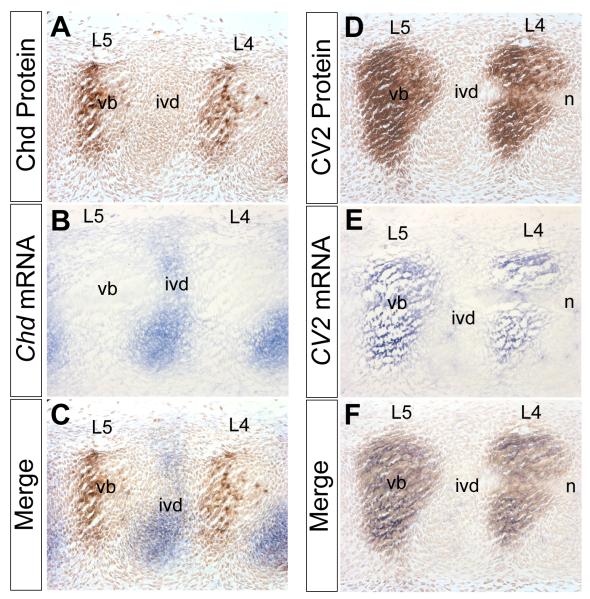
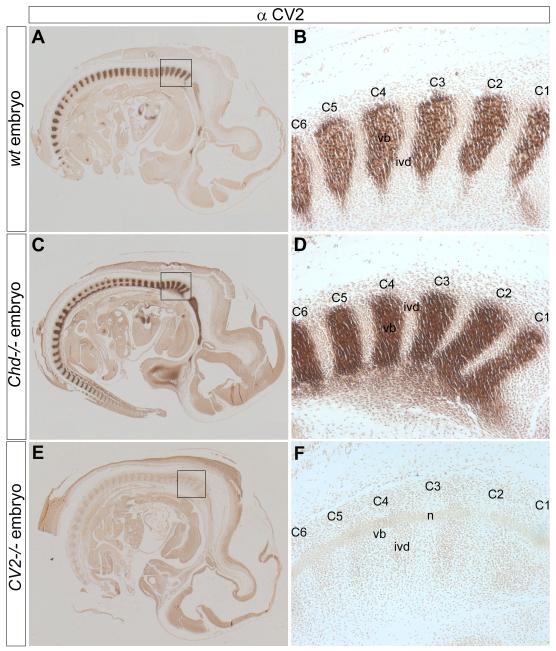
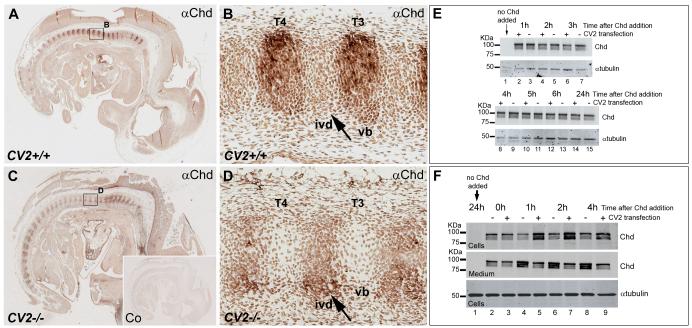
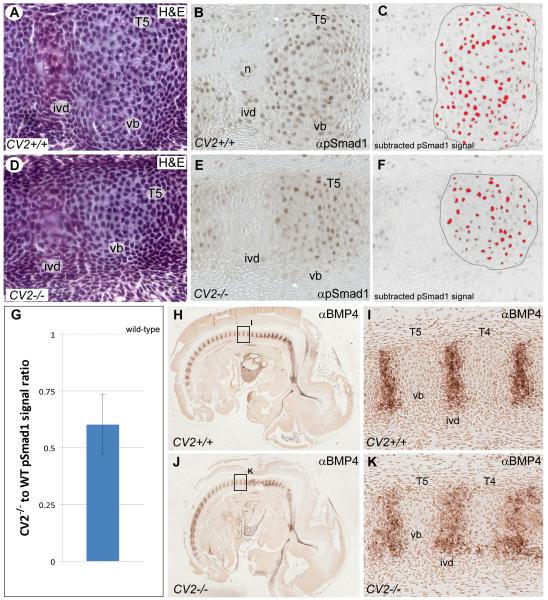
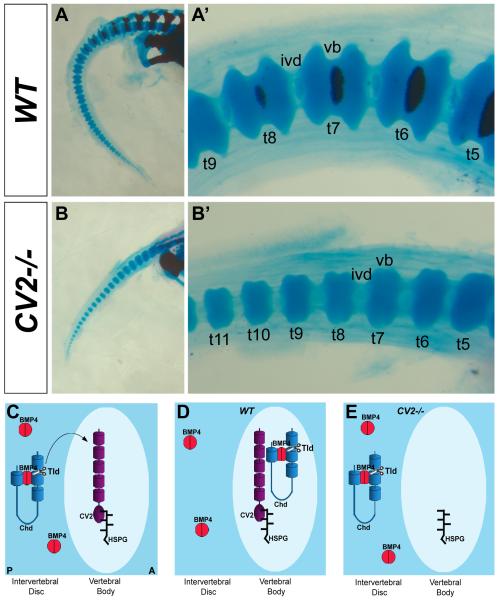
Bone morphogenetic proteins (BMPs), as well as the BMP-binding molecules Chordin (Chd), Crossveinless-2 (CV2) and Twisted Gastrulation (Tsg), are essential for axial skeletal development in the mouse embryo. We previously reported a strong genetic interaction between CV2 and Tsg and proposed a role for this interaction in the shaping of the BMP morphogenetic field during vertebral development. In the present study we investigated the roles of CV2 and Chd in the formation of the vertebral morphogenetic field. We performed immunostainings for CV2 and Chd protein on wild-type, CV2(-/-) or Chd(-/-) mouse embryo sections at the stage of onset of the vertebral phenotypes. By comparing mRNA and protein localizations we found that CV2 does not diffuse away from its place of synthesis, the vertebral body. The most interesting finding of this study was that Chd synthesized in the intervertebral disc accumulates in the vertebral body. This relocalization does not take place in CV2(-/-) mutants. Instead, Chd was found to accumulate at its site of synthesis in CV2(-/-) embryos. These results indicate a CV2-dependent flow of Chd protein from the intervertebral disc to the vertebral body. Smad1/5/8 phosphorylation was decreased in CV2(-/-)vertebral bodies. This impaired BMP signaling may result from the decreased levels of Chd/BMP complexes diffusing from the intervertebral region. The data indicate a role for CV2 and Chd in the establishment of the vertebral morphogenetic field through the long-range relocalization of Chd/BMP complexes. The results may have general implications for the formation of embryonic organ-forming morphogenetic fields.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Development of the vertebral morphogenetic field in the mouse: interactions between Crossveinless-2 and Twisted Gastrulation.Dev Biol. 2008 Nov 1;323(1):6-18. doi: 10.1016/j.ydbio.2008.08.019. Epub 2008 Aug 29. Dev Biol. 2008. PMID: 18789316 Free PMC article.
-
Crossveinless-2 Is a BMP feedback inhibitor that binds Chordin/BMP to regulate Xenopus embryonic patterning.Dev Cell. 2008 Aug;15(2):248-60. doi: 10.1016/j.devcel.2008.06.013. Dev Cell. 2008. PMID: 18694564 Free PMC article.
-
Binding between Crossveinless-2 and Chordin von Willebrand factor type C domains promotes BMP signaling by blocking Chordin activity.PLoS One. 2010 Sep 23;5(9):e12846. doi: 10.1371/journal.pone.0012846. PLoS One. 2010. PMID: 20886103 Free PMC article.
-
The Chordin Morphogenetic Pathway.Curr Top Dev Biol. 2016;116:231-45. doi: 10.1016/bs.ctdb.2015.10.003. Epub 2016 Jan 7. Curr Top Dev Biol. 2016. PMID: 26970622 Review.
-
The role of chordin fragments generated by partial tolloid cleavage in regulating BMP activity.Biochem Soc Trans. 2015 Oct;43(5):795-800. doi: 10.1042/BST20150071. Biochem Soc Trans. 2015. PMID: 26517884 Free PMC article. Review.
Cited by
-
Premature Vertebral Mineralization in hmx1-Mutant Zebrafish.Cells. 2022 Mar 24;11(7):1088. doi: 10.3390/cells11071088. Cells. 2022. PMID: 35406651 Free PMC article.
-
Scale invariance of BMP signaling gradients in zebrafish.Sci Rep. 2019 Apr 1;9(1):5440. doi: 10.1038/s41598-019-41840-8. Sci Rep. 2019. PMID: 30932076 Free PMC article.
-
Multistep molecular mechanism for bone morphogenetic protein extracellular transport in the Drosophila embryo.Proc Natl Acad Sci U S A. 2012 Jul 10;109(28):11222-7. doi: 10.1073/pnas.1202781109. Epub 2012 Jun 25. Proc Natl Acad Sci U S A. 2012. PMID: 22733779 Free PMC article.
-
Fine-tuned shuttles for bone morphogenetic proteins.Curr Opin Genet Dev. 2013 Aug;23(4):374-84. doi: 10.1016/j.gde.2013.04.012. Epub 2013 Jun 2. Curr Opin Genet Dev. 2013. PMID: 23735641 Free PMC article. Review.
-
Development of the axial skeleton and intervertebral disc.Curr Top Dev Biol. 2019;133:49-90. doi: 10.1016/bs.ctdb.2018.11.018. Epub 2019 Jan 3. Curr Top Dev Biol. 2019. PMID: 30902259 Free PMC article. Review.
References
-
- Bachiller D, Klingensmith J, Kemp C, Belo JA, Anderson RM, May SR, McMahon JA, McMahon AP, Harland RM, Rossant J, De Robertis EM. The organizer factors Chordin and Noggin are required for mouse forebrain development. Nature. 2000;403:658–661. - PubMed
-
- Bachiller D, Klingensmith J, Shneyder N, Tran U, Anderson R, Rossant J, De Robertis EM. The role of Chordin/BMP signals in mammalian pharyngeal development and DiGeorge syndrome. Development. 2003;130:3567–3578. - PubMed
-
- Bagnall KM, Higgins SJ, Sanders EJ. The contribution made by a single somite to the vertebral column: experimental evidence in support of resegmentation using the chick-quail chimaera model. Development. 1988;103:69–85. - PubMed
-
- Ben-Zvi D, Shilo BZ, Fainsod A, Barkai N. Scaling of the BMP activation gradient in Xenopus embryos. Nature. 2008;453:1205–1211. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
