

Cyclin-dependent kinase-mediated phosphorylation plays a critical role in the oncogenic functions of PELP1
- PMID: 20807815
- PMCID: PMC3058498
- DOI: 10.1158/0008-5472.CAN-10-0628
Cyclin-dependent kinase-mediated phosphorylation plays a critical role in the oncogenic functions of PELP1
Abstract
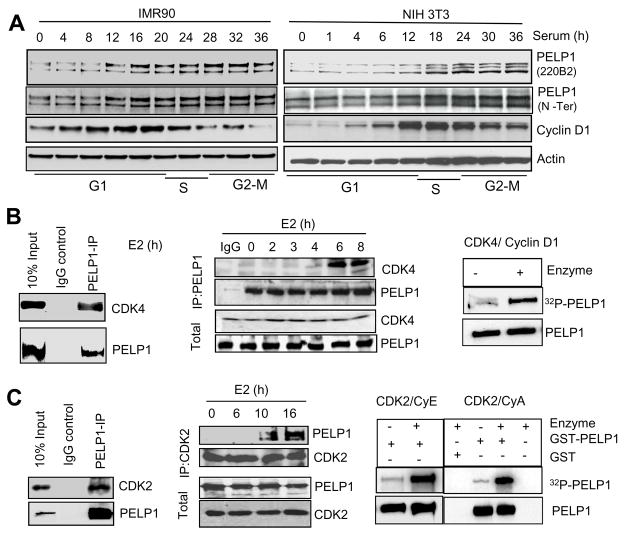
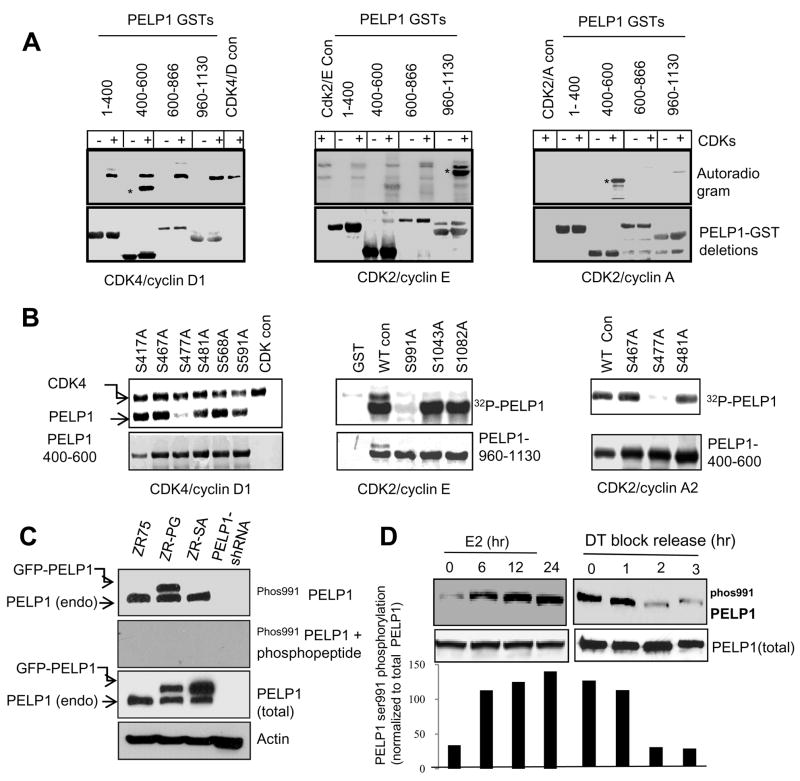
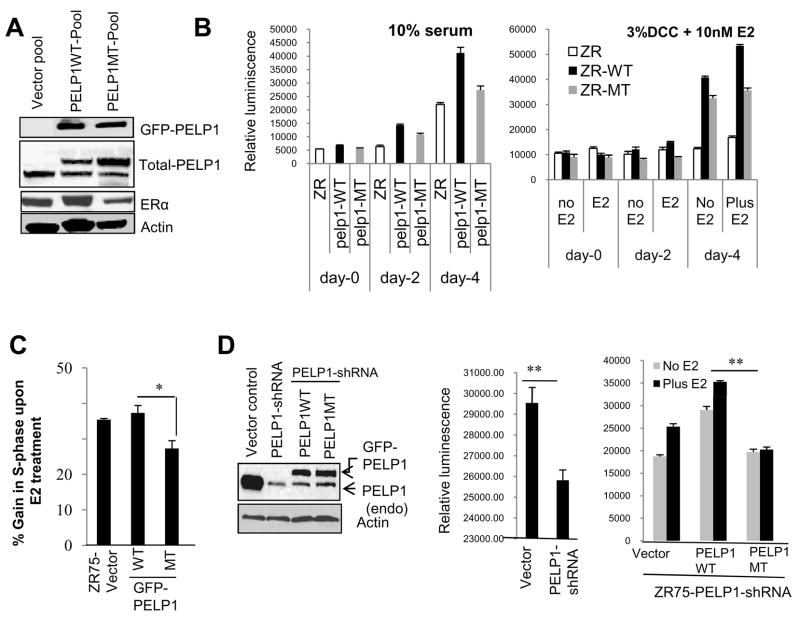
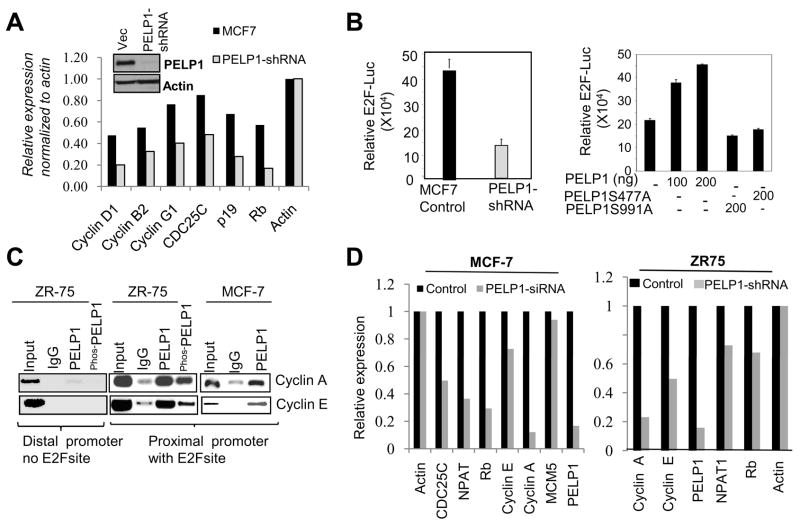
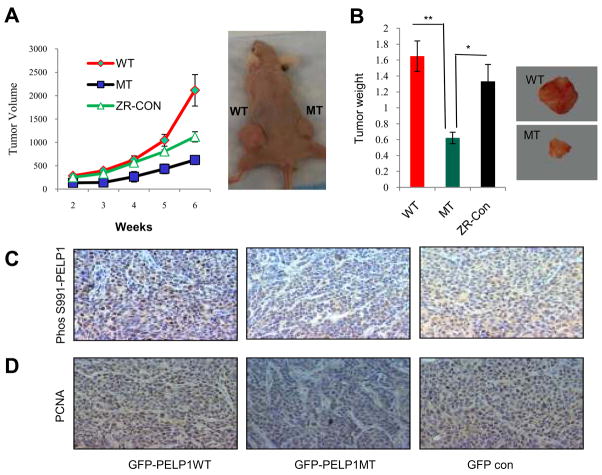
Estrogen receptor (ER) signaling plays an important role in breast cancer progression, and ER functions are influenced by coregulatory proteins. PELP1 (proline-, glutamic acid-, and leucine-rich protein 1) is a nuclear receptor coregulator that plays an important role in ER signaling. Its expression is deregulated in hormonal cancers. We identified PELP1 as a novel cyclin-dependent kinase (CDK) substrate. Using site-directed mutagenesis and in vitro kinase assays, we identified Ser(477) and Ser(991) of PELP1 as CDK phosphorylation sites. Using the PELP1 Ser(991) phospho-specific antibody, we show that PELP1 is hyperphosphorylated during cell cycle progression. Model cells stably expressing the PELP1 mutant that lack CDK sites had defects in estradiol (E2)-mediated cell cycle progression and significantly affected PELP1-mediated oncogenic functions in vivo. Mechanistic studies showed that PELP1 modulates transcription factor E2F1 transactivation functions, that PELP1 is recruited to pRb/E2F target genes, and that PELP1 facilitates ER signaling cross talk with cell cycle machinery. We conclude that PELP1 is a novel substrate of interphase CDKs and that its phosphorylation is important for the proper function of PELP1 in modulating hormone-driven cell cycle progression and also for optimal E2F transactivation function. Because the expression of both PELP1 and CDKs is deregulated in breast tumors, CDK-PELP1 interactions will have implications in breast cancer progression.
©2010 AACR.
Figures
Similar articles
-
Oncogenic potential of the nuclear receptor coregulator proline-, glutamic acid-, leucine-rich protein 1/modulator of the nongenomic actions of the estrogen receptor.Cancer Res. 2007 Jun 1;67(11):5505-12. doi: 10.1158/0008-5472.CAN-06-3647. Cancer Res. 2007. PMID: 17545633 Free PMC article.
-
Growth factor regulation of estrogen receptor coregulator PELP1 functions via Protein Kinase A pathway.Mol Cancer Res. 2008 May;6(5):851-61. doi: 10.1158/1541-7786.MCR-07-2030. Mol Cancer Res. 2008. PMID: 18505929 Free PMC article.
-
Modulation of in situ estrogen synthesis by proline-, glutamic acid-, and leucine-rich protein-1: potential estrogen receptor autocrine signaling loop in breast cancer cells.Mol Endocrinol. 2008 Mar;22(3):649-64. doi: 10.1210/me.2007-0350. Epub 2007 Dec 13. Mol Endocrinol. 2008. PMID: 18079323 Free PMC article.
-
PELP1: Structure, biological function and clinical significance.Gene. 2016 Jul 1;585(1):128-134. doi: 10.1016/j.gene.2016.03.017. Epub 2016 Mar 18. Gene. 2016. PMID: 26997260 Free PMC article. Review.
-
Comprehensive analysis of recent biochemical and biologic findings regarding a newly discovered protein-PELP1/MNAR.Clin Exp Metastasis. 2006;23(1):1-7. doi: 10.1007/s10585-006-9019-9. Epub 2006 Jul 7. Clin Exp Metastasis. 2006. PMID: 16826428 Review.
Cited by
-
Minireview: Deciphering the Cellular Functions of PELP1.Mol Endocrinol. 2015 Sep;29(9):1222-9. doi: 10.1210/me.2015-1049. Epub 2015 Jul 9. Mol Endocrinol. 2015. PMID: 26158753 Free PMC article. Review. No abstract available.
-
PELP1/SRC-3-dependent regulation of metabolic PFKFB kinases drives therapy resistant ER+ breast cancer.Oncogene. 2021 Jun;40(25):4384-4397. doi: 10.1038/s41388-021-01871-w. Epub 2021 Jun 8. Oncogene. 2021. PMID: 34103681 Free PMC article.
-
Cryo-EM reveals the architecture of the PELP1-WDR18 molecular scaffold.Nat Commun. 2022 Nov 9;13(1):6783. doi: 10.1038/s41467-022-34610-0. Nat Commun. 2022. PMID: 36351913 Free PMC article.
-
Significance of PELP1/HDAC2/miR-200 regulatory network in EMT and metastasis of breast cancer.Oncogene. 2014 Jul 10;33(28):3707-16. doi: 10.1038/onc.2013.332. Epub 2013 Aug 26. Oncogene. 2014. PMID: 23975430 Free PMC article.
-
Minireview: Recent advances in extranuclear steroid receptor actions.Endocrinology. 2011 Dec;152(12):4489-95. doi: 10.1210/en.2011-1470. Epub 2011 Oct 25. Endocrinology. 2011. PMID: 22028449 Free PMC article. Review.
References
-
- Sherr CJ. Cancer cell cycles. Science. 1996;274:1672–7. - PubMed
-
- Berthet C, Klarmann KD, Hilton MB, Suh HC, Keller JR, Kiyokawa H, et al. Combined loss of Cdk2 and Cdk4 results in embryonic lethality and Rb hypophosphorylation. Dev Cell. 2006;10:563–73. - PubMed
-
- Foster JS, Henley DC, Ahamed S, Wimalasena J. Estrogens and cell-cycle regulation in breast cancer. Trends Endocrinol Metab. 2001;12:320–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
