

Glycosaminoglycan sulphation affects the seeded misfolding of a mutant prion protein
- PMID: 20808809
- PMCID: PMC2925953
- DOI: 10.1371/journal.pone.0012351
Glycosaminoglycan sulphation affects the seeded misfolding of a mutant prion protein
Abstract
Background: The accumulation of protease resistant conformers of the prion protein (PrP(res)) is a key pathological feature of prion diseases. Polyanions, including RNA and glycosaminoglycans have been identified as factors that contribute to the propagation, transmission and pathogenesis of prion disease. Recent studies have suggested that the contribution of these cofactors to prion propagation may be species specific.
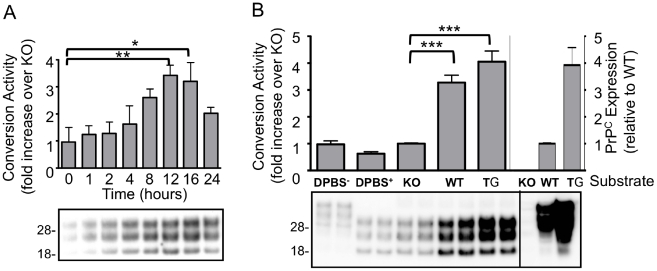
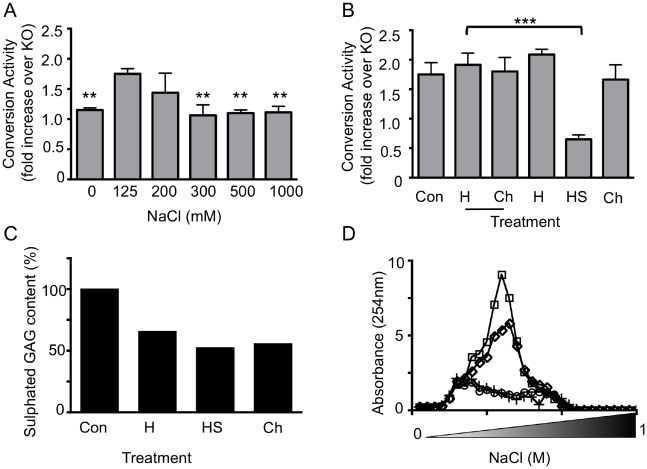
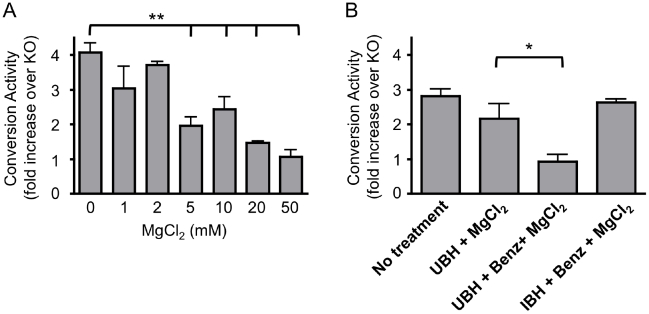
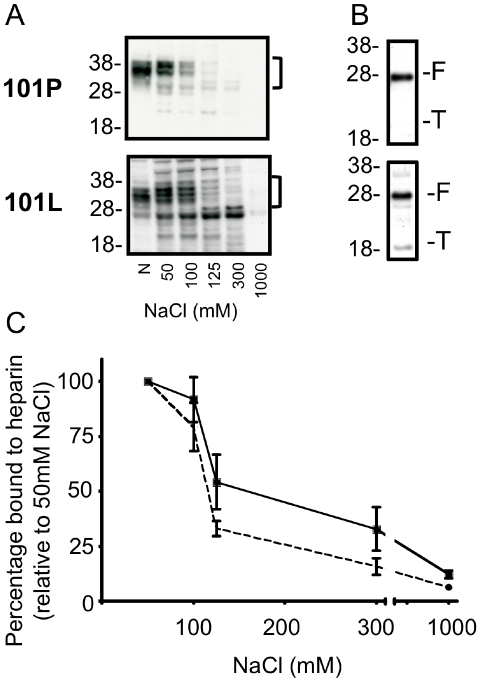
Methodology/principal finding: In this study a cell-free assay was used to investigate the molecular basis of polyanion stimulated PrP(res) formation using brain tissue or cell line derived murine PrP. Enzymatic depletion of endogenous nucleic acids or heparan sulphate (HS) from the PrP(C) substrate was found to specifically prevent PrP(res) formation seeded by mouse derived PrP(Sc). Modification of the negative charge afforded by the sulphation of glycosaminoglycans increased the ability of a familial PrP mutant to act as a substrate for PrP(res) formation, while having no effect on PrP(res) formed by wildtype PrP. This difference may be due to the observed differences in the binding of wild type and mutant PrP for glycosaminoglycans.
Conclusions/significance: Cofactor requirements for PrP(res) formation are host species and prion strain specific and affected by disease associated mutations of the prion protein. This may explain both species and strain dependent propagation characteristics and provide insights into the underlying mechanisms of familial prion disease. It further highlights the challenge of designing effective therapeutics against a disease which effects a range of mammalian species, caused by range of aetiologies and prion strains.
Conflict of interest statement
Figures
Similar articles
-
Glycosaminoglycan sulfation determines the biochemical properties of prion protein aggregates.Glycobiology. 2015 Jul;25(7):745-55. doi: 10.1093/glycob/cwv014. Epub 2015 Feb 20. Glycobiology. 2015. PMID: 25701659
-
Requirements for mutant and wild-type prion protein misfolding in vitro.Biochemistry. 2015 Feb 10;54(5):1180-7. doi: 10.1021/bi501495j. Epub 2015 Jan 22. Biochemistry. 2015. PMID: 25584902 Free PMC article.
-
In vitro amplification of scrapie and chronic wasting disease PrP(res) using baculovirus-expressed recombinant PrP as substrate.Prion. 2014;8(6):393-403. doi: 10.4161/19336896.2014.983753. Prion. 2014. PMID: 25495764 Free PMC article.
-
Prion neurotoxicity: insights from prion protein mutants.Curr Issues Mol Biol. 2010;12(2):51-61. Epub 2009 Sep 18. Curr Issues Mol Biol. 2010. PMID: 19767650 Free PMC article. Review.
-
PrP interactions with nucleic acids and glycosaminoglycans in function and disease.Front Biosci (Landmark Ed). 2010 Jan 1;15(1):132-50. doi: 10.2741/3611. Front Biosci (Landmark Ed). 2010. PMID: 20036811 Review.
Cited by
-
Prion subcellular fractionation reveals infectivity spectrum, with a high titre-low PrPres level disparity.Mol Neurodegener. 2012 Apr 26;7:18. doi: 10.1186/1750-1326-7-18. Mol Neurodegener. 2012. PMID: 22534096 Free PMC article.
-
Organoids for modeling prion diseases.Cell Tissue Res. 2023 Apr;392(1):97-111. doi: 10.1007/s00441-022-03589-x. Epub 2022 Jan 28. Cell Tissue Res. 2023. PMID: 35088182 Free PMC article. Review.
-
Heparan Sulfate and Heparin Promote Faithful Prion Replication in Vitro by Binding to Normal and Abnormal Prion Proteins in Protein Misfolding Cyclic Amplification.J Biol Chem. 2016 Dec 16;291(51):26478-26486. doi: 10.1074/jbc.M116.745851. Epub 2016 Nov 7. J Biol Chem. 2016. PMID: 27821590 Free PMC article.
-
The prion protein preference of sporadic Creutzfeldt-Jakob disease subtypes.J Biol Chem. 2012 Oct 19;287(43):36465-72. doi: 10.1074/jbc.M112.368803. Epub 2012 Aug 28. J Biol Chem. 2012. PMID: 22930754 Free PMC article. Clinical Trial.
-
Heparan sulfate S-domains and extracellular sulfatases (Sulfs): their possible roles in protein aggregation diseases.Glycoconj J. 2018 Aug;35(4):387-396. doi: 10.1007/s10719-018-9833-8. Epub 2018 Jul 12. Glycoconj J. 2018. PMID: 30003471 Review.
References
-
- Collins SJ, Lawson VA, Masters CL. Transmissible spongiform encephalopathies. Lancet. 2004;363:51–61. - PubMed
-
- Bueler H, Aguzzi A, Sailer A, Greiner RA, Autenried P, et al. Mice devoid of PrP are resistant to scrapie. Cell. 1993;73:1339–1347. - PubMed
-
- Prusiner SB. Novel proteinaceous infectious particles cause scrapie. Science. 1982;216:136–144. - PubMed
-
- Kocisko DA, Come JH, Priola SA, Chesebro B, Raymond GJ, et al. Cell-free formation of protease-resistant prion protein. Nature. 1994;370:471–474. - PubMed
-
- Lucassen R, Nishina K, Supattapone S. In vitro amplification of protease-resistant prion protein requires free sulfhydryl groups. Biochemistry. 2003;42:4127–4135. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
