

A Toxoplasma MORN1 null mutant undergoes repeated divisions but is defective in basal assembly, apicoplast division and cytokinesis
- PMID: 20808817
- PMCID: PMC2924399
- DOI: 10.1371/journal.pone.0012302
A Toxoplasma MORN1 null mutant undergoes repeated divisions but is defective in basal assembly, apicoplast division and cytokinesis
Abstract
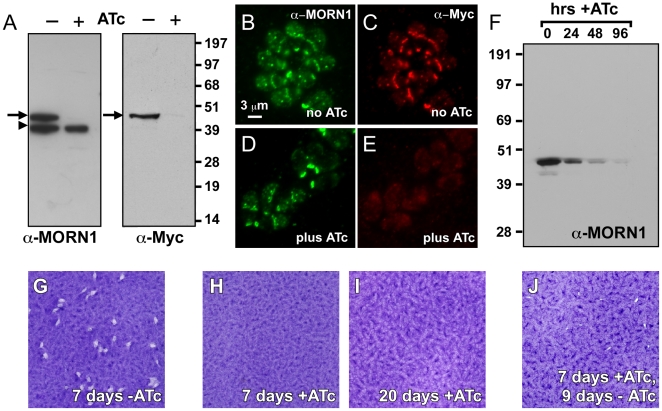
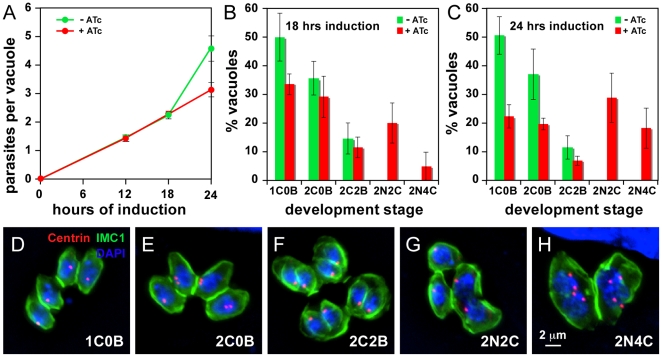
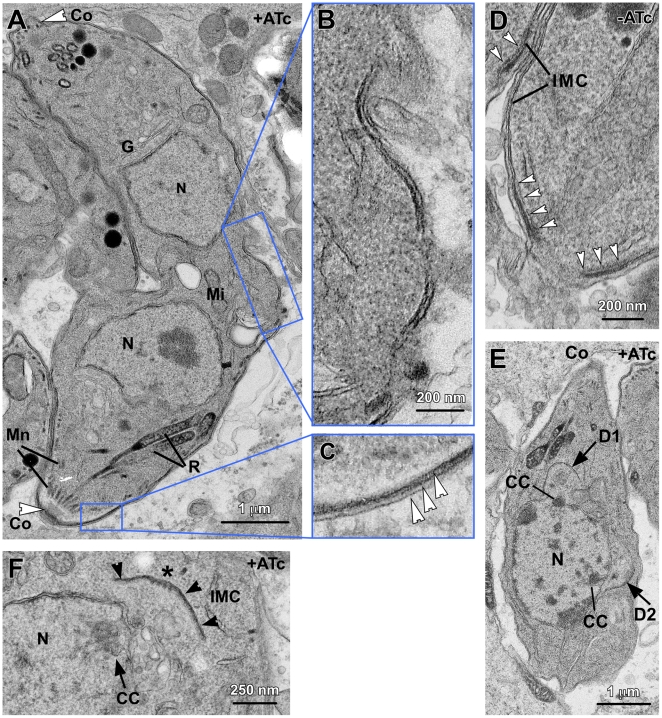
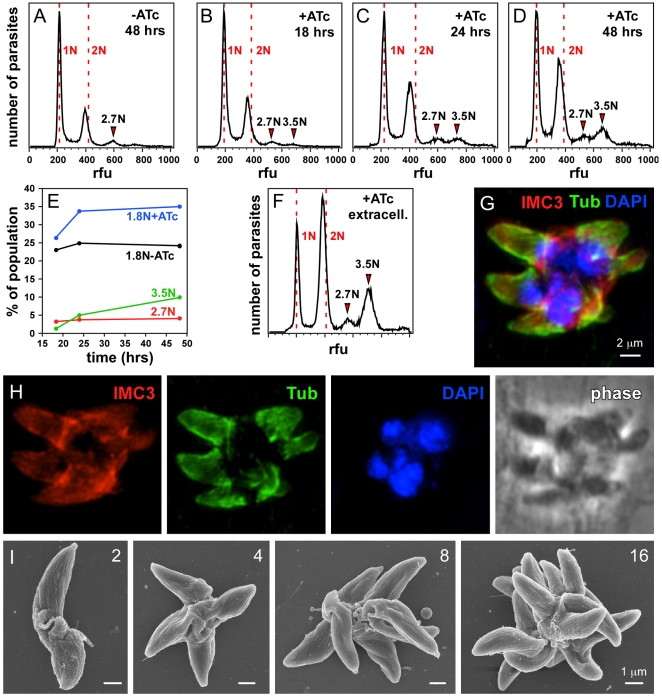
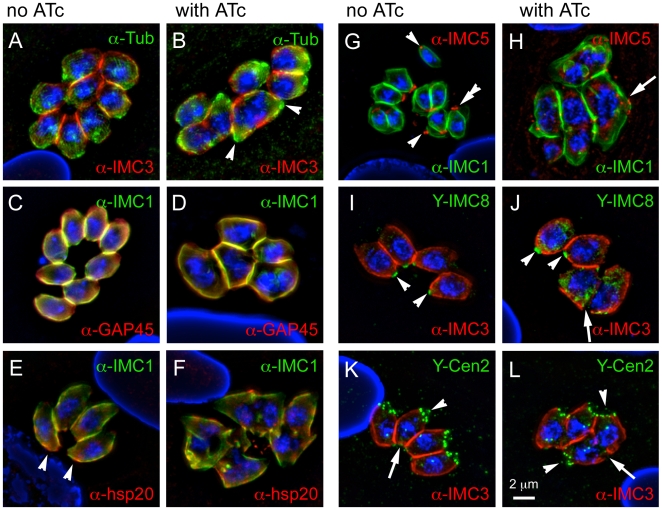
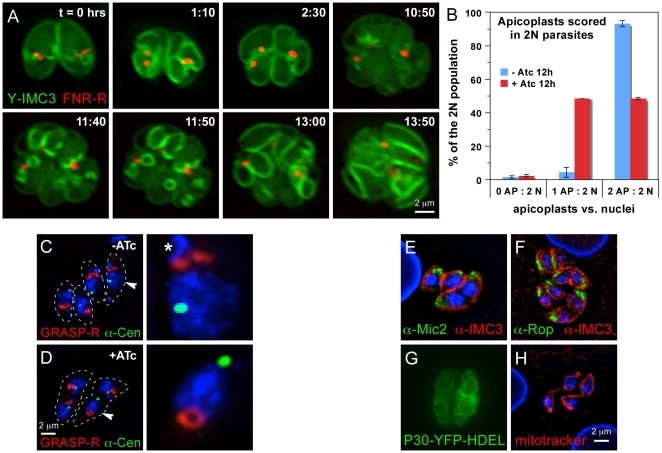
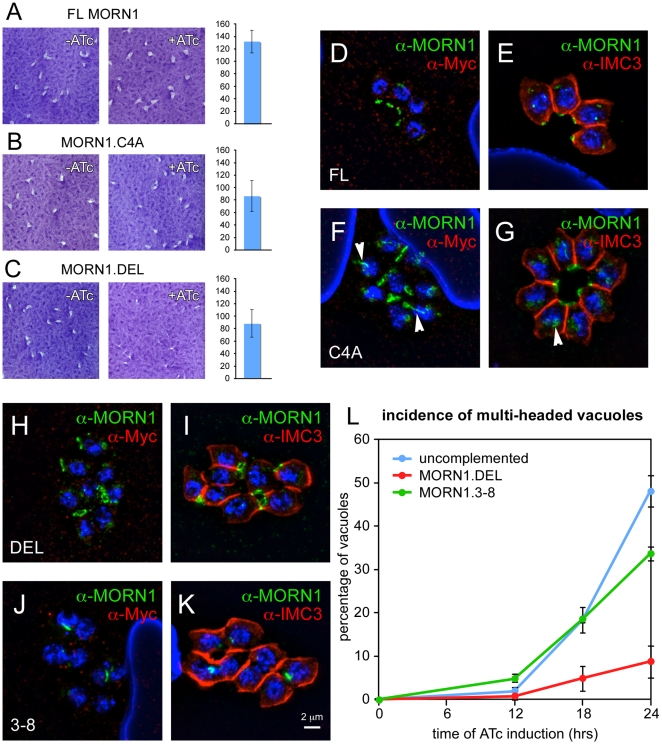
The membrane occupation and recognition nexus protein 1 (MORN1) is highly conserved among apicomplexan parasites and is associated with several structures that have a role in cell division. Here we dissected the role of MORN1 using the relatively simple budding process of Toxoplasma gondii as a model. Ablation of MORN1 in a conditional null mutant resulted in pronounced defects suggesting a central role for MORN1 in apicoplast segregation and in daughter cell budding. Lack of MORN1 resulted in double-headed parasites. These Janus-headed parasites form two complete apical complexes but fail to assemble a basal complex. Moreover, these parasites were capable of undergoing several more budding rounds resulting in the formation of up to 16-headed parasites conjoined at the basal end. Despite this segregation defect, the mother's cytoskeleton was completely disassembled in every budding round. Overall this argues that successful completion of the budding is not required for cell cycle progression. None of the known basal complex components, including a set of recently identified inner membrane complex (IMC) proteins, localized correctly in these multi-headed parasites. These data suggest that MORN1 is essential for assembly of the basal complex, and that lack of the basal complex abolishes the contractile capacity assigned to the basal complex late in daughter formation. Consistent with this hypothesis we observe that MORN1 mutants fail to efficiently constrict and divide the apicoplast. We used the null background provided by the mutant to dissect the function of subdomains of the MORN1 protein. This demonstrated that deletion of a single MORN domain already prevented the function of MORN1 whereas a critical role for the short linker between MORN domains 6 and 7 was identified. In conclusion, MORN1 is required for basal complex assembly and loss of MORN1 results in defects in apicoplast division and daughter segregation.
Conflict of interest statement
Figures
Similar articles
-
A MORN-repeat protein is a dynamic component of the Toxoplasma gondii cell division apparatus.J Cell Sci. 2006 Jun 1;119(Pt 11):2236-45. doi: 10.1242/jcs.02949. Epub 2006 May 9. J Cell Sci. 2006. PMID: 16684814
-
A MORN1-associated HAD phosphatase in the basal complex is essential for Toxoplasma gondii daughter budding.Cell Microbiol. 2016 Aug;18(8):1153-71. doi: 10.1111/cmi.12574. Epub 2016 Mar 9. Cell Microbiol. 2016. PMID: 26840427 Free PMC article.
-
Targeted proteomic dissection of Toxoplasma cytoskeleton sub-compartments using MORN1.Cytoskeleton (Hoboken). 2012 Dec;69(12):1069-85. doi: 10.1002/cm.21077. Epub 2012 Oct 11. Cytoskeleton (Hoboken). 2012. PMID: 23027733 Free PMC article.
-
Organelle Dynamics in Apicomplexan Parasites.mBio. 2021 Aug 31;12(4):e0140921. doi: 10.1128/mBio.01409-21. Epub 2021 Aug 24. mBio. 2021. PMID: 34425697 Free PMC article. Review.
-
Cytoskeleton assembly in Toxoplasma gondii cell division.Int Rev Cell Mol Biol. 2012;298:1-31. doi: 10.1016/B978-0-12-394309-5.00001-8. Int Rev Cell Mol Biol. 2012. PMID: 22878103 Free PMC article. Review.
Cited by
-
Ciprofloxacin Derivatives Affect Parasite Cell Division and Increase the Survival of Mice Infected with Toxoplasma gondii.PLoS One. 2015 May 7;10(5):e0125705. doi: 10.1371/journal.pone.0125705. eCollection 2015. PLoS One. 2015. PMID: 25950173 Free PMC article.
-
The apical annuli of Toxoplasma gondii are composed of coiled-coil and signalling proteins embedded in the inner membrane complex sutures.Cell Microbiol. 2020 Jan;22(1):e13112. doi: 10.1111/cmi.13112. Epub 2019 Sep 10. Cell Microbiol. 2020. PMID: 31470470 Free PMC article.
-
The SUN-like protein TgSLP1 is essential for nuclear division in the apicomplexan parasite Toxoplasma gondii.J Cell Sci. 2023 Nov 1;136(21):jcs260337. doi: 10.1242/jcs.260337. Epub 2023 Oct 30. J Cell Sci. 2023. PMID: 37815466 Free PMC article.
-
Plasticity between MyoC- and MyoA-glideosomes: an example of functional compensation in Toxoplasma gondii invasion.PLoS Pathog. 2014 Nov 13;10(10):e1004504. doi: 10.1371/journal.ppat.1004504. eCollection 2014 Oct. PLoS Pathog. 2014. PMID: 25393004 Free PMC article.
-
Pellicle formation in the malaria parasite.J Cell Sci. 2016 Feb 15;129(4):673-80. doi: 10.1242/jcs.181230. Epub 2016 Jan 13. J Cell Sci. 2016. PMID: 26763910 Free PMC article.
References
-
- Montoya JG, Liesenfeld O. Toxoplasmosis. Lancet. 2004;363:1965–1976. - PubMed
-
- Sheffield HG, Melton ML. The fine structure and reproduction of Toxoplasma gondii. J Parasitol. 1968;54:209–226. - PubMed
-
- Gubbels MJ, Vaishnava S, Boot N, Dubremetz JF, Striepen B. A MORN-repeat protein is a dynamic component of the Toxoplasma gondii cell division apparatus. J Cell Sci. 2006;119:2236–2245. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
