

DNA-triggered aggregation of copper, zinc superoxide dismutase in the presence of ascorbate
- PMID: 20808835
- PMCID: PMC2924893
- DOI: 10.1371/journal.pone.0012328
DNA-triggered aggregation of copper, zinc superoxide dismutase in the presence of ascorbate
Abstract
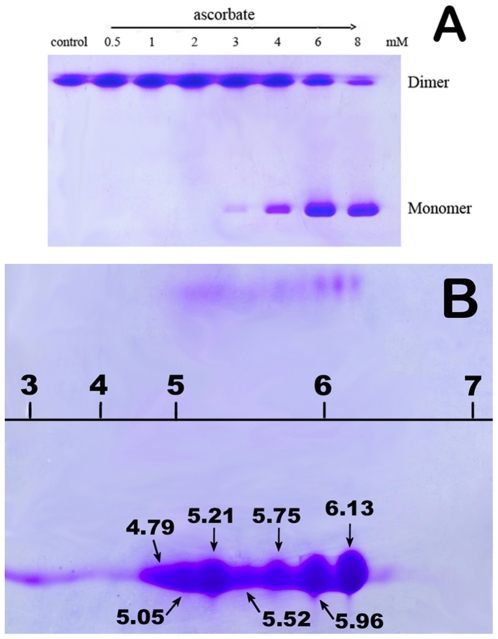
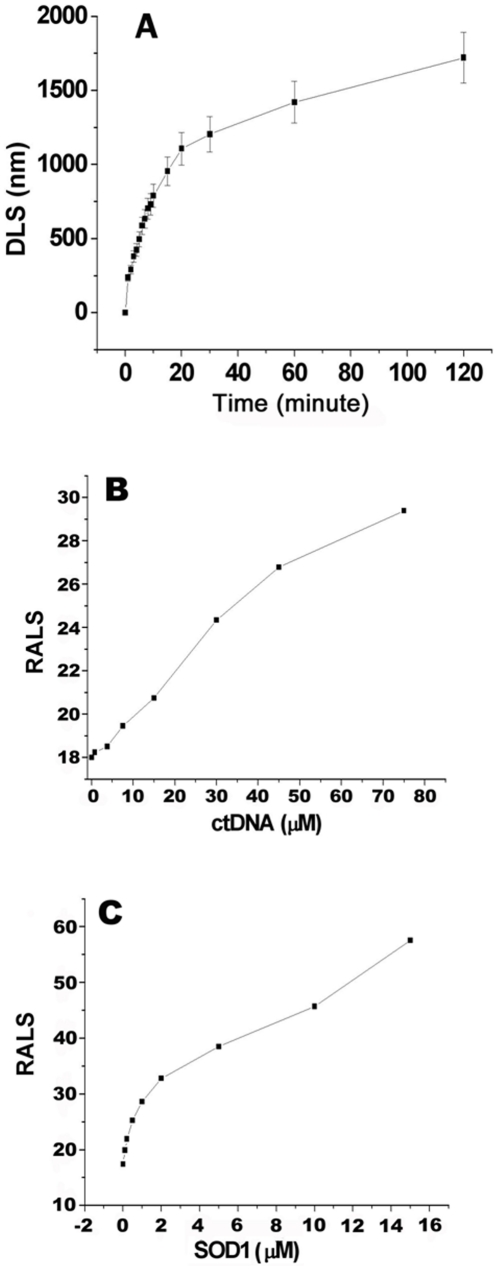
The oxidative damage hypothesis proposed for the function gain of copper, zinc superoxide dismutase (SOD1) maintains that both mutant and wild-type (WT) SOD1 catalyze reactions with abnormal substrates that damage cellular components critical for viability of the affected cells. However, whether the oxidative damage of SOD1 is involved in the formation of aggregates rich in SOD1 or not remains elusive. Here, we sought to explore the oxidative aggregation of WT SOD1 exposed to environments containing both ascorbate (Asc) and DNA under neutral conditions. The results showed that the WT SOD1 protein was oxidized in the presence of Asc. The oxidation results in the higher affinity of the modified protein for DNA than that of the unmodified protein. The oxidized SOD1 was observed to be more prone to aggregation than the WT SOD1, and the addition of DNA can significantly accelerate the oxidative aggregation. Moreover, a reasonable relationship can be found between the oxidation, increased hydrophobicity, and aggregation of SOD1 in the presence of DNA. The crucial step in aggregation is neutralization of the positive charges on some SOD1 surfaces by DNA binding. This study might be crucial for understanding molecular forces driving the protein aggregation.
Conflict of interest statement
Figures




Similar articles
-
Cupric ions induce the oxidation and trigger the aggregation of human superoxide dismutase 1.PLoS One. 2013 Jun 3;8(6):e65287. doi: 10.1371/journal.pone.0065287. Print 2013. PLoS One. 2013. PMID: 23755211 Free PMC article.
-
DNA is a template for accelerating the aggregation of copper, zinc superoxide dismutase.Biochemistry. 2007 May 22;46(20):5911-23. doi: 10.1021/bi062234m. Epub 2007 May 1. Biochemistry. 2007. PMID: 17469801
-
Polyanion binding accelerates the formation of stable and low-toxic aggregates of ALS-linked SOD1 mutant A4V.Proteins. 2014 Dec;82(12):3356-72. doi: 10.1002/prot.24691. Epub 2014 Oct 1. Proteins. 2014. PMID: 25220364
-
Mutant SOD1 instability: implications for toxicity in amyotrophic lateral sclerosis.Neurodegener Dis. 2005;2(3-4):115-27. doi: 10.1159/000089616. Neurodegener Dis. 2005. PMID: 16909016 Review.
-
Structure, folding, and misfolding of Cu,Zn superoxide dismutase in amyotrophic lateral sclerosis.Biochim Biophys Acta. 2006 Nov-Dec;1762(11-12):1025-37. doi: 10.1016/j.bbadis.2006.05.004. Epub 2006 May 22. Biochim Biophys Acta. 2006. PMID: 16814528 Review.
Cited by
-
DNA Facilitates Oligomerization and Prevents Aggregation via DNA Networks.Biophys J. 2020 Jan 7;118(1):162-171. doi: 10.1016/j.bpj.2019.11.022. Epub 2019 Nov 23. Biophys J. 2020. PMID: 31839258 Free PMC article.
-
SAXS Examinations of the Redox-Dependent Formation of a DNA-SOD1 Complex.Int J Mol Sci. 2022 Oct 21;23(20):12673. doi: 10.3390/ijms232012673. Int J Mol Sci. 2022. PMID: 36293533 Free PMC article.
References
-
- Fridovich I. Superoxide dismutases. Annu Rev Biochem. 1975;44:147–159. - PubMed
-
- Kinnula VL, Crapo J. Superoxide dismutase in malihnant cells and human tumors. Free Radic Biol Med. 2004;36:718–744. - PubMed
-
- Ezzi SA, Urashitani M, Julien J-P. Wild-type superoxide dismutase acquires binding and toxic properties of ALS-linked mutant forms through oxidation. J Neurochem. 2007;102:170–178. - PubMed
-
- Jaarsma D, Haasdijk ED, Grashorn JAC, Hawkins RW, Verspaget HW, et al. Human Cu/Zn superoxide dismutase (SOD1) overexpression in mice cause mitochondrial vacuolization, axonal degradation, and premature motor neuron death and accelerates motor neuron disease in mice expressing a familial amyotrophic lateral sclerosis mutant SOD1. Neurobiol Dis. 2000;7:623–643. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
