

Ancient protostome origin of chemosensory ionotropic glutamate receptors and the evolution of insect taste and olfaction
- PMID: 20808886
- PMCID: PMC2924276
- DOI: 10.1371/journal.pgen.1001064
Ancient protostome origin of chemosensory ionotropic glutamate receptors and the evolution of insect taste and olfaction
Abstract
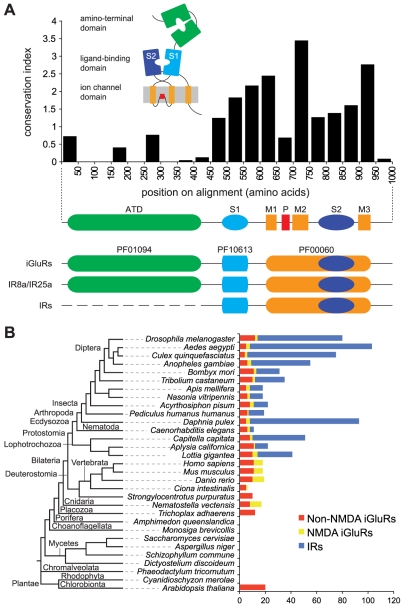
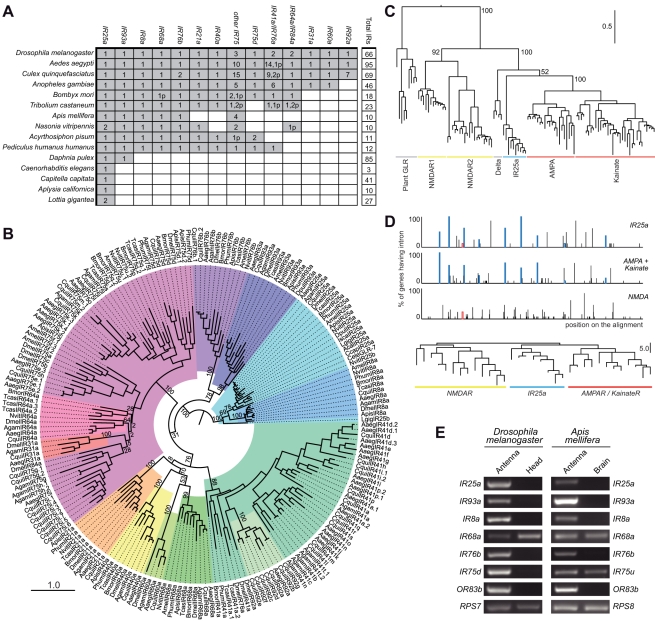
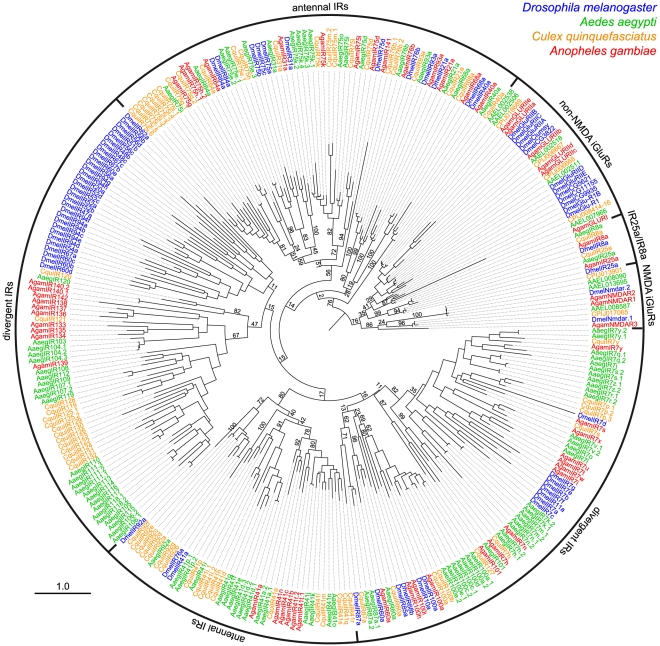
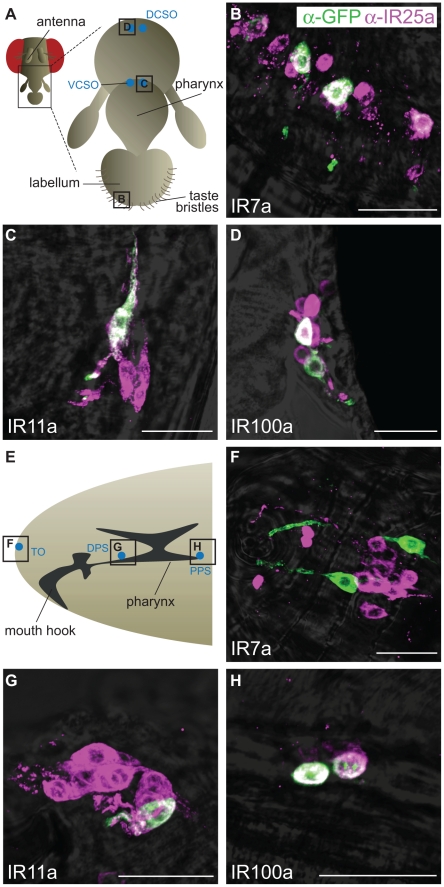
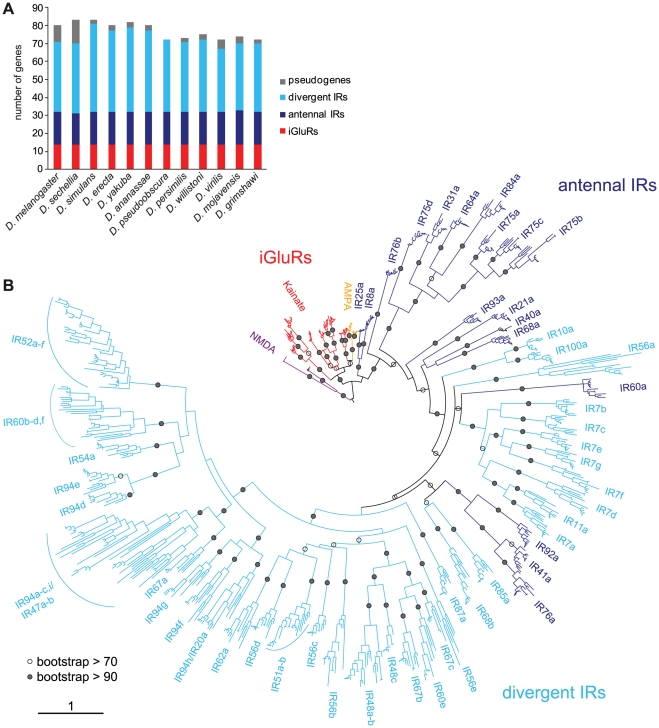
Ionotropic glutamate receptors (iGluRs) are a highly conserved family of ligand-gated ion channels present in animals, plants, and bacteria, which are best characterized for their roles in synaptic communication in vertebrate nervous systems. A variant subfamily of iGluRs, the Ionotropic Receptors (IRs), was recently identified as a new class of olfactory receptors in the fruit fly, Drosophila melanogaster, hinting at a broader function of this ion channel family in detection of environmental, as well as intercellular, chemical signals. Here, we investigate the origin and evolution of IRs by comprehensive evolutionary genomics and in situ expression analysis. In marked contrast to the insect-specific Odorant Receptor family, we show that IRs are expressed in olfactory organs across Protostomia--a major branch of the animal kingdom that encompasses arthropods, nematodes, and molluscs--indicating that they represent an ancestral protostome chemosensory receptor family. Two subfamilies of IRs are distinguished: conserved "antennal IRs," which likely define the first olfactory receptor family of insects, and species-specific "divergent IRs," which are expressed in peripheral and internal gustatory neurons, implicating this family in taste and food assessment. Comparative analysis of drosophilid IRs reveals the selective forces that have shaped the repertoires in flies with distinct chemosensory preferences. Examination of IR gene structure and genomic distribution suggests both non-allelic homologous recombination and retroposition contributed to the expansion of this multigene family. Together, these findings lay a foundation for functional analysis of these receptors in both neurobiological and evolutionary studies. Furthermore, this work identifies novel targets for manipulating chemosensory-driven behaviours of agricultural pests and disease vectors.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
-
- Gereau RW, Swanson GT. The Glutamate Receptors. Totowa, N.J.: Humana Press; 2008. p. xi, 576.
-
- Armstrong N, Sun Y, Chen GQ, Gouaux E. Structure of a glutamate-receptor ligand-binding core in complex with kainate. Nature. 1998;395:913–917. - PubMed
-
- Kuner T, Seeburg PH, Guy HR. A common architecture for K+ channels and ionotropic glutamate receptors? Trends Neurosci. 2003;26:27–32. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
