

Molecular mechanisms leading to null-protein product from retinoschisin (RS1) signal-sequence mutants in X-linked retinoschisis (XLRS) disease
- PMID: 20809529
- PMCID: PMC2991635
- DOI: 10.1002/humu.21350
Molecular mechanisms leading to null-protein product from retinoschisin (RS1) signal-sequence mutants in X-linked retinoschisis (XLRS) disease
Abstract
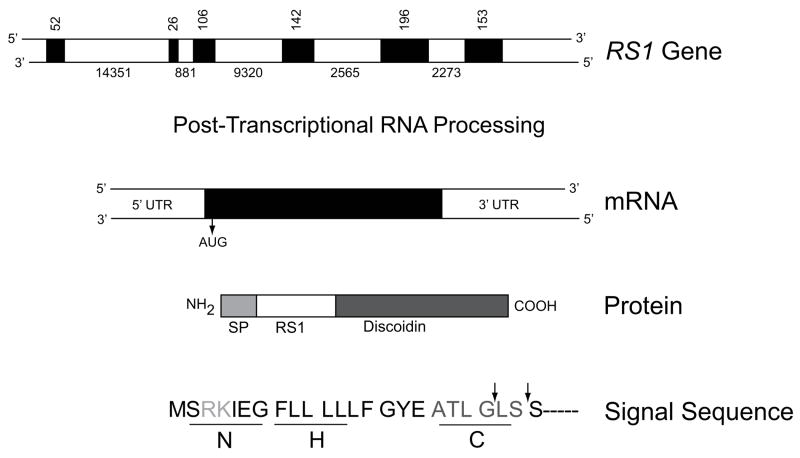
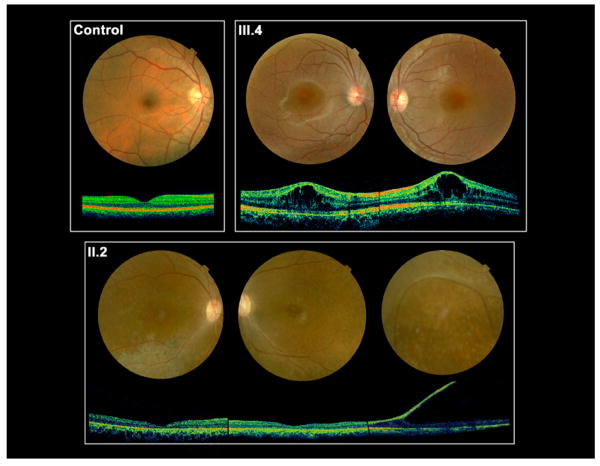
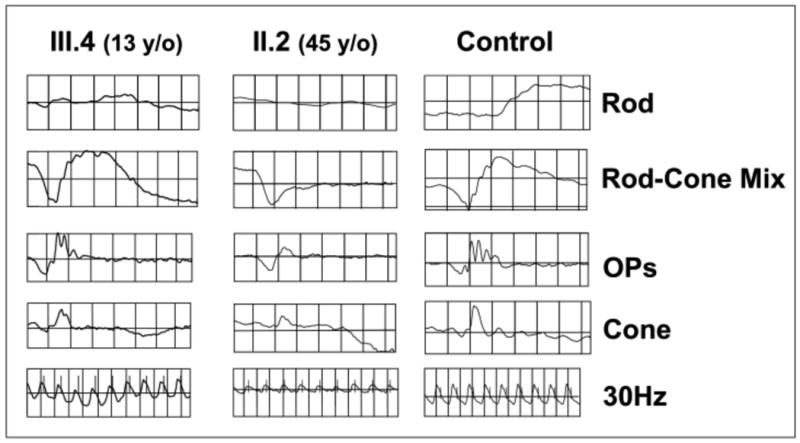
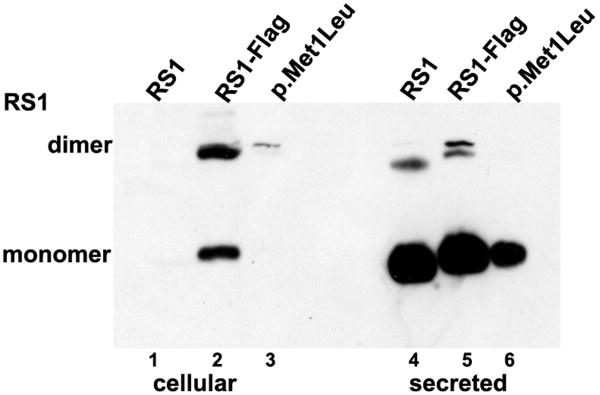
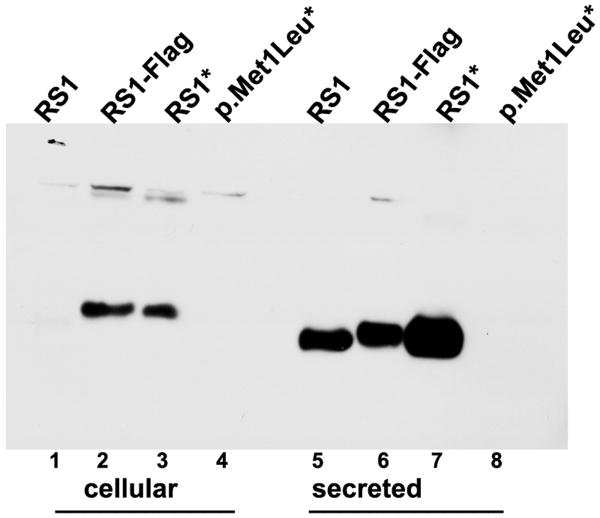
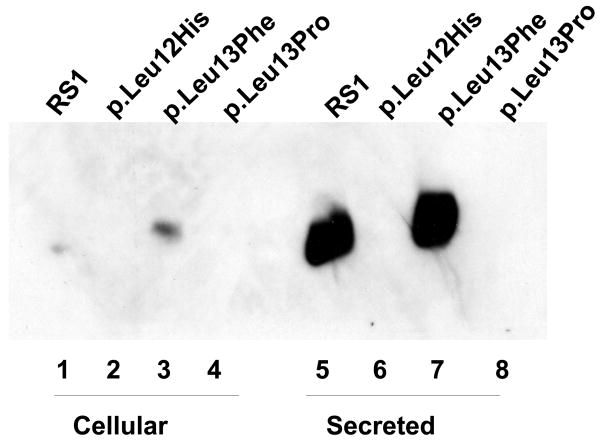
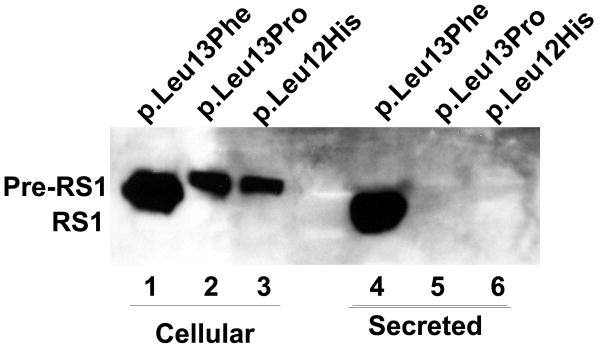
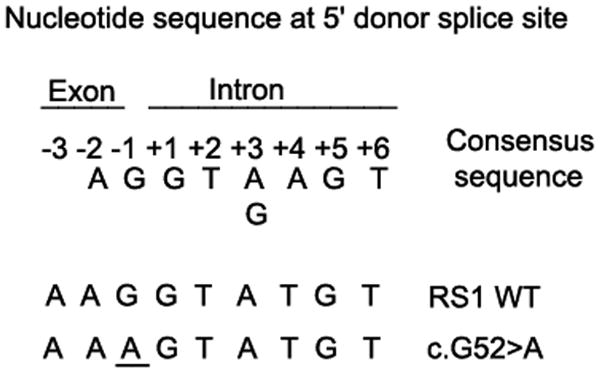
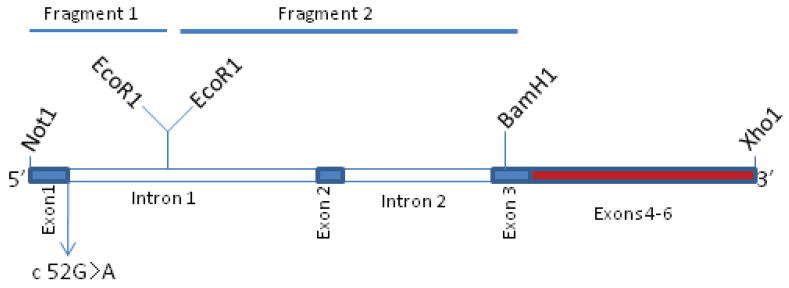
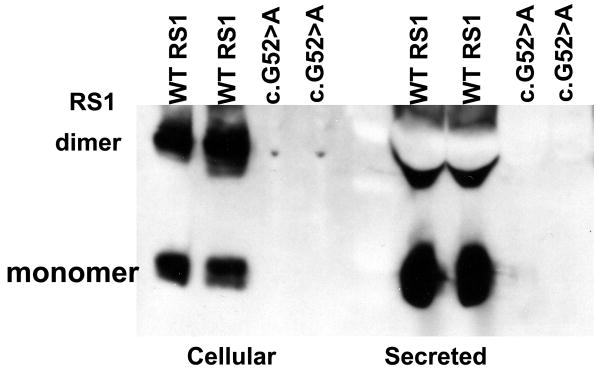
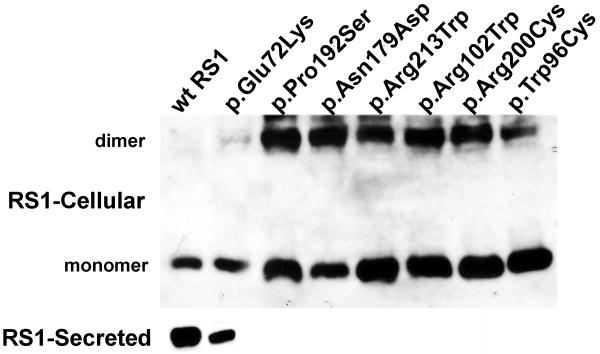
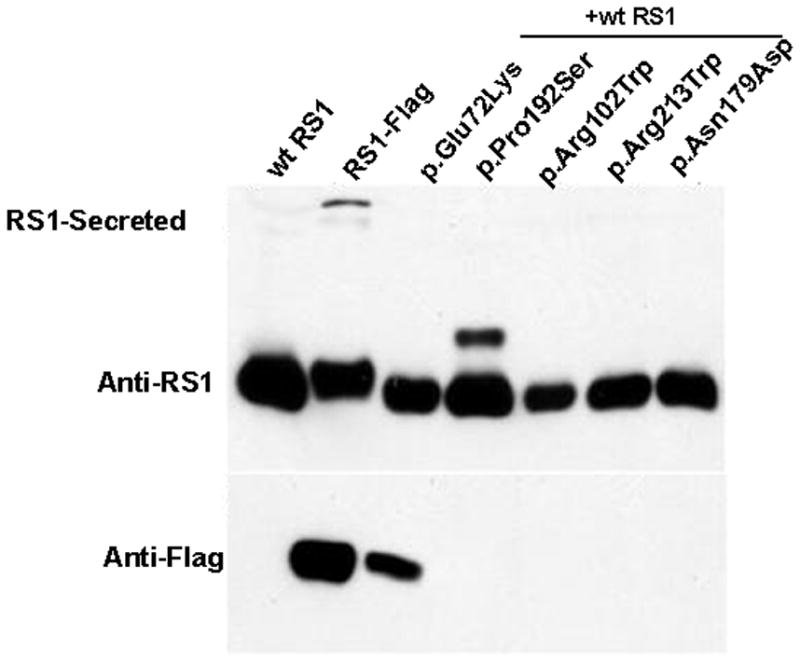
Retinoschisin (RS1) is a cell-surface adhesion molecule expressed by photoreceptor and bipolar cells of the retina. The 24-kDa protein encodes two conserved sequence motifs: the initial signal sequence targets the protein for secretion while the larger discoidin domain is implicated in cell adhesion. RS1 helps to maintain the structural organization of the retinal cell layers and promotes visual signal transduction. RS1 gene mutations cause X-linked retinoschisis disease (XLRS) in males, characterized by early-onset central vision loss. We analyzed the biochemical consequences of several RS1 signal-sequence mutants (c.1A>T, c.35T>A, c.38T>C, and c.52G>A) found in our subjects. Expression analysis in COS-7 cells demonstrates that these mutations affect RS1 biosynthesis and result in an RS1 null phenotype by several different mechanisms. By comparison, discoidin-domain mutations generally lead to nonfunctional conformational variants that remain trapped inside the cell. XLRS disease has a broad heterogeneity in general, but subjects with the RS1 null-protein signal-sequence mutations are on the more severe end of the clinical phenotype. Results from the signal-sequence mutants are discussed in the context of the discoidin-domain mutations, clinical phenotypes, genotype-phenotype correlations, and implications for RS1 gene replacement therapy.
This article is a US Government work and, as such, is in the public domain in the United States of America. Published in 2010 by Wiley-Liss, Inc.
Figures
Similar articles
-
Of men and mice: Human X-linked retinoschisis and fidelity in mouse modeling.Prog Retin Eye Res. 2022 Mar;87:100999. doi: 10.1016/j.preteyeres.2021.100999. Epub 2021 Aug 11. Prog Retin Eye Res. 2022. PMID: 34390869 Review.
-
Null retinoschisin-protein expression from an RS1 c354del1-ins18 mutation causing progressive and severe XLRS in a cross-sectional family study.Invest Ophthalmol Vis Sci. 2009 Nov;50(11):5375-83. doi: 10.1167/iovs.09-3839. Epub 2009 May 27. Invest Ophthalmol Vis Sci. 2009. PMID: 19474399 Free PMC article.
-
X-linked juvenile retinoschisis: clinical diagnosis, genetic analysis, and molecular mechanisms.Prog Retin Eye Res. 2012 May;31(3):195-212. doi: 10.1016/j.preteyeres.2011.12.002. Epub 2012 Jan 3. Prog Retin Eye Res. 2012. PMID: 22245536 Free PMC article. Review.
-
Coexpression and interaction of wild-type and missense RS1 mutants associated with X-linked retinoschisis: its relevance to gene therapy.Invest Ophthalmol Vis Sci. 2007 Jun;48(6):2491-7. doi: 10.1167/iovs.06-1465. Invest Ophthalmol Vis Sci. 2007. PMID: 17525175
-
RS1, a discoidin domain-containing retinal cell adhesion protein associated with X-linked retinoschisis, exists as a novel disulfide-linked octamer.J Biol Chem. 2005 Mar 18;280(11):10721-30. doi: 10.1074/jbc.M413117200. Epub 2005 Jan 11. J Biol Chem. 2005. PMID: 15644328
Cited by
-
A novel deletion mutation in RS1 gene caused X-linked juvenile retinoschisis in a Chinese family.Eye (Lond). 2014 Nov;28(11):1364-9. doi: 10.1038/eye.2014.196. Epub 2014 Aug 29. Eye (Lond). 2014. PMID: 25168411 Free PMC article.
-
Early Developmental Characteristics and Features of a Three-Dimensional Retinal Organoid Model of X-Linked Juvenile Retinoschisis.Int J Mol Sci. 2024 Jul 27;25(15):8203. doi: 10.3390/ijms25158203. Int J Mol Sci. 2024. PMID: 39125773 Free PMC article.
-
Treatment-Emergent Adverse Events in Gene Therapy Trials for Inherited Retinal Diseases: A Narrative Review.Ophthalmol Ther. 2020 Dec;9(4):709-724. doi: 10.1007/s40123-020-00287-1. Epub 2020 Aug 1. Ophthalmol Ther. 2020. PMID: 32740739 Free PMC article. Review.
-
Molecular modeling indicates distinct classes of missense variants with mild and severe XLRS phenotypes.Hum Mol Genet. 2013 Dec 1;22(23):4756-67. doi: 10.1093/hmg/ddt329. Epub 2013 Jul 11. Hum Mol Genet. 2013. PMID: 23847049 Free PMC article.
-
Phenotypic expression of X-linked retinoschisis in Chinese families with mutations in the RS1 gene.Doc Ophthalmol. 2011 Aug;123(1):21-7. doi: 10.1007/s10633-011-9278-x. Epub 2011 Jun 24. Doc Ophthalmol. 2011. PMID: 21701876
References
-
- Akli S, Chelly J, Mezard C, Gandy S, Kahn A, Poenaru L. A “G” to “A” mutation at position -1 of a 5′ splice site in a late infantile form of Tay-Sachs disease. J Biol Chem. 1990;265(13):7324–30. - PubMed
-
- Andrews LG, Markert ML. Exon skipping in purine nucleoside phosphorylase mRNA processing leading to severe immunodeficiency. J Biol Chem. 1992;267(11):7834–8. - PubMed
-
- Antoun A, Pavlov MY, Lovmar M, Ehrenberg M. How initiation factors maximize the accuracy of tRNA selection in initiation of bacterial protein synthesis. Mol Cell. 2006;23(2):183–93. - PubMed
-
- Bradshaw K, Allen L, Trump D, Hardcastle A, George N, Moore A. A comparison of ERG abnormalities in XLRS and XLCSNB. Doc Ophthalmol. 2004;108(2):135–45. - PubMed
-
- Bradshaw K, George N, Moore A, Trump D. Mutations of the XLRS1 gene cause abnormalities of photoreceptor as well as inner retinal responses of the ERG. Doc Ophthalmol. 1999;98(2):153–73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
