

Enrichment and differential targeting of complexins 3 and 4 in ribbon-containing sensory neurons during zebrafish development
- PMID: 20809954
- PMCID: PMC2941751
- DOI: 10.1186/1749-8104-5-24
Enrichment and differential targeting of complexins 3 and 4 in ribbon-containing sensory neurons during zebrafish development
Abstract
Background: In sensory systems with broad bandwidths, polarized receptor cells utilize highly specialized organelles in their apical and basolateral compartments to transduce and ultimately transmit signals to the rest of the nervous system. While progress has been made in elucidating the assembly of the transduction apparatus, the development of synaptic ribbon-containing terminals remains poorly understood. To begin to delineate the targeting of the exocytotic machinery specifically in ribbon-containing neurons, we have examined the expression of complexins 3 and 4 in the zebrafish visual and acousticolateral systems during the first week of development.
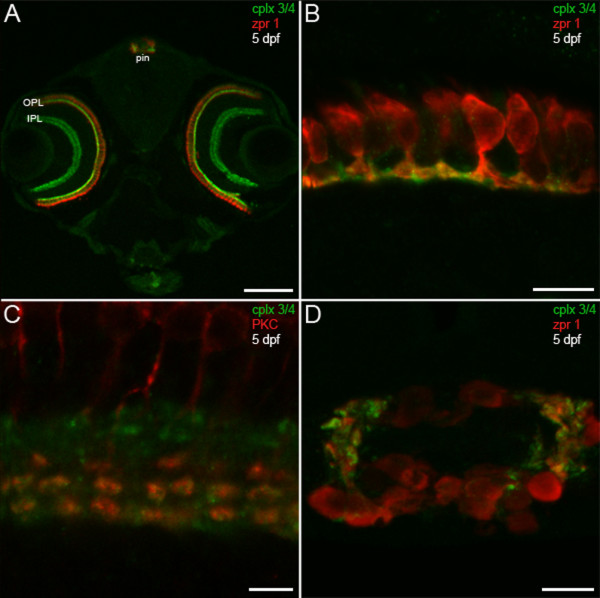
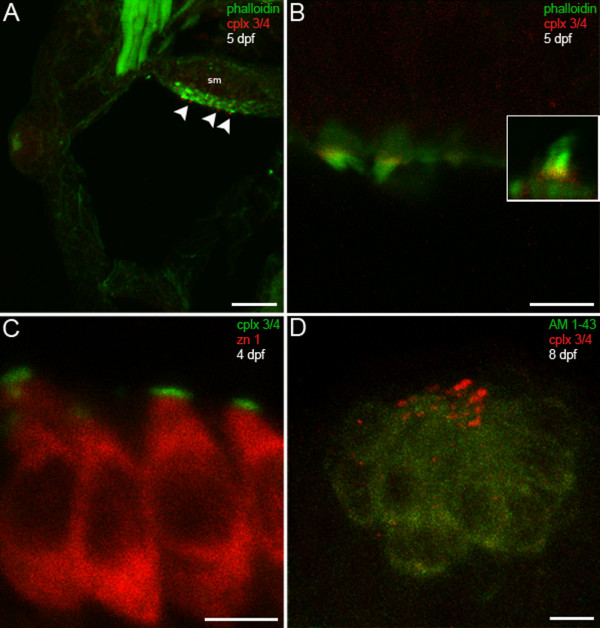
Results: We have identified five members of the complexin 3/4 subfamily in zebrafish that show 50 to 75% amino acid identity with mammalian complexins 3 and 4. Utilizing a polyclonal antibody that recognizes all five orthologs, we demonstrate that these proteins are enriched in ribbon-containing sensory neurons. Complexin 3/4 is rapidly targeted to presynaptic terminals in the pineal organ and retina concomitantly with RIBEYE b, a component of ribbons. In hair cells of the inner ear and lateral line, however, complexin 3/4 immunoreactivity clusters on the apical surfaces of hair cells, among their stereocilia, rather than along the basolateral plasma membrane with RIBEYE b. A complexin 4a-specific antibody selectively labels the presynaptic terminals of visual system ribbon-containing neurons.
Conclusions: These results provide evidence for the concurrent transport and/or assembly of multiple components of the active zone in developing ribbon terminals. Members of the complexin 3/4 subfamily are enriched in these terminals in the visual system and in hair bundles of the acousticolateral system, suggesting that these proteins are differentially targeted and may have multiple roles in ribbon-containing sensory neurons.
Figures









Similar articles
-
Ribeye is required for presynaptic Ca(V)1.3a channel localization and afferent innervation of sensory hair cells.Development. 2011 Apr;138(7):1309-19. doi: 10.1242/dev.059451. Epub 2011 Feb 24. Development. 2011. PMID: 21350006 Free PMC article.
-
Characterization of Ribeye subunits in zebrafish hair cells reveals that exogenous Ribeye B-domain and CtBP1 localize to the basal ends of synaptic ribbons.PLoS One. 2014 Sep 10;9(9):e107256. doi: 10.1371/journal.pone.0107256. eCollection 2014. PLoS One. 2014. PMID: 25208216 Free PMC article.
-
Presynaptic Nrxn3 is essential for ribbon-synapse maturation in hair cells.Development. 2024 Oct 1;151(19):dev202723. doi: 10.1242/dev.202723. Epub 2024 Oct 10. Development. 2024. PMID: 39254120 Free PMC article.
-
Dual use of the transcriptional repressor (CtBP2)/ribbon synapse (RIBEYE) gene: how prevalent are multifunctional genes?Trends Neurosci. 2001 Oct;24(10):555-7. doi: 10.1016/s0166-2236(00)01894-4. Trends Neurosci. 2001. PMID: 11576649 Review.
-
The making of synaptic ribbons: how they are built and what they do.Neuroscientist. 2009 Dec;15(6):611-24. doi: 10.1177/1073858409340253. Neuroscientist. 2009. PMID: 19700740 Review.
Cited by
-
Development and maintenance of vision's first synapse.Dev Biol. 2021 Aug;476:218-239. doi: 10.1016/j.ydbio.2021.04.001. Epub 2021 Apr 10. Dev Biol. 2021. PMID: 33848537 Free PMC article. Review.
-
Transcriptome analysis in mice treated with vigabatrin identifies dysregulation of genes associated with retinal signaling circuitry.Epilepsy Res. 2020 Oct;166:106395. doi: 10.1016/j.eplepsyres.2020.106395. Epub 2020 Jun 20. Epilepsy Res. 2020. PMID: 32679486 Free PMC article.
-
The dynamic architecture of photoreceptor ribbon synapses: cytoskeletal, extracellular matrix, and intramembrane proteins.Vis Neurosci. 2011 Nov;28(6):453-71. doi: 10.1017/S0952523811000356. Vis Neurosci. 2011. PMID: 22192503 Free PMC article. Review.
-
Functional roles of complexin in neurotransmitter release at ribbon synapses of mouse retinal bipolar neurons.J Neurosci. 2015 Mar 4;35(9):4065-70. doi: 10.1523/JNEUROSCI.2703-14.2015. J Neurosci. 2015. PMID: 25740533 Free PMC article.
-
Stabilization of spontaneous neurotransmitter release at ribbon synapses by ribbon-specific subtypes of complexin.J Neurosci. 2013 May 8;33(19):8216-26. doi: 10.1523/JNEUROSCI.1280-12.2013. J Neurosci. 2013. PMID: 23658160 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
