

Oxygen sensing in Drosophila: multiple isoforms of the prolyl hydroxylase fatiga have different capacity to regulate HIFalpha/Sima
- PMID: 20811646
- PMCID: PMC2928329
- DOI: 10.1371/journal.pone.0012390
Oxygen sensing in Drosophila: multiple isoforms of the prolyl hydroxylase fatiga have different capacity to regulate HIFalpha/Sima
Abstract
Background: The Hypoxia Inducible Factor (HIF) mediates cellular adaptations to low oxygen. Prolyl-4-hydroxylases are oxygen sensors that hydroxylate the HIF alpha-subunit, promoting its proteasomal degradation in normoxia. Three HIF-prolyl hydroxylases, encoded by independent genes, PHD1, PHD2, and PHD3, occur in mammals. PHD2, the longest PHD isoform includes a MYND domain, whose biochemical function is unclear. PHD2 and PHD3 genes are induced in hypoxia to shut down HIF dependent transcription upon reoxygenation, while expression of PHD1 is oxygen-independent. The physiologic significance of the diversity of the PHD oxygen sensors is intriguing.
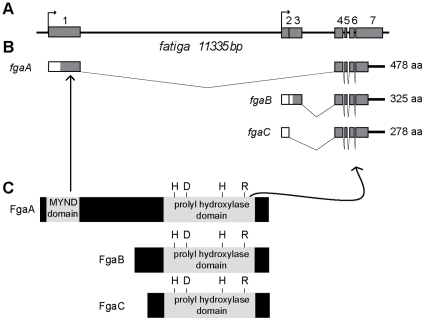
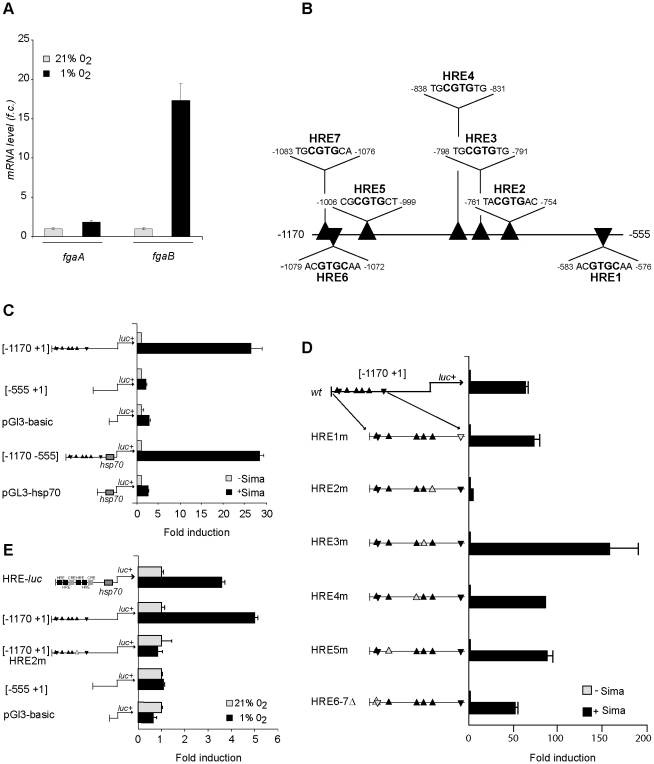
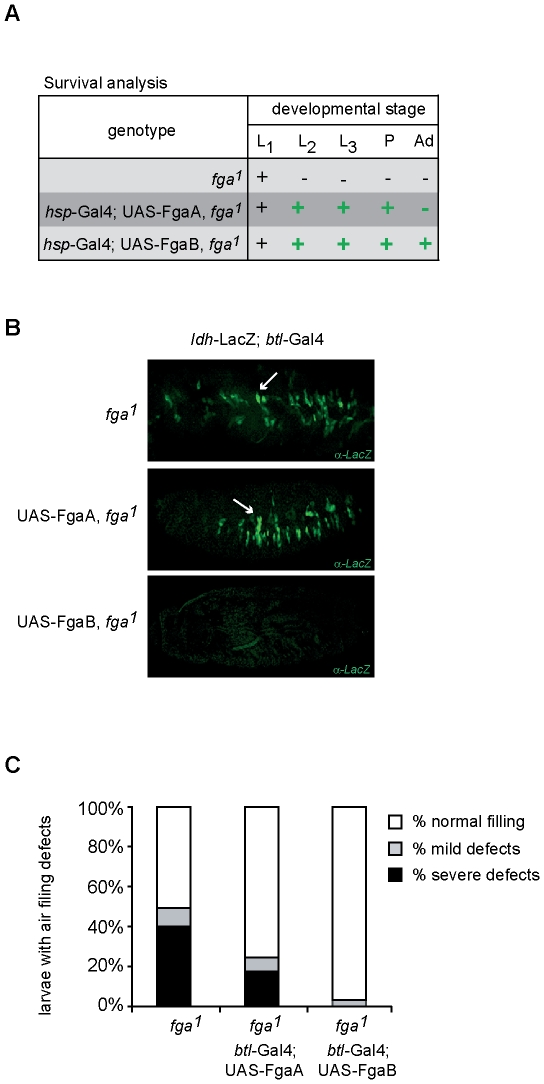
Methodology and principal findings: We have analyzed the Drosophila PHD locus, fatiga, which encodes 3 isoforms, FgaA, FgaB and FgaC that are originated through a combination of alternative initiation of transcription and alternative splicing. FgaA includes a MYND domain and is homologous to PHD2, while FgaB and FgaC are shorter isoforms most similar to PHD3. Through a combination of genetic experiments in vivo and molecular analyses in cell culture, we show that fgaB but not fgaA is induced in hypoxia, in a Sima-dependent manner, through a HIF-Responsive Element localized in the first intron of fgaA. The regulatory capacity of FgaB is stronger than that of FgaA, as complete reversion of fga loss-of-function phenotypes is observed upon transgenic expression of the former, and only partial rescue occurs after expression of the latter.
Conclusions and significance: Diversity of PHD isoforms is a conserved feature in evolution. As in mammals, there are hypoxia-inducible and non-inducible Drosophila PHDs, and a fly isoform including a MYND domain co-exists with isoforms lacking this domain. Our results suggest that the isoform devoid of a MYND domain has stronger regulatory capacity than that including this domain.
Conflict of interest statement
Figures

References
-
- Bruick RK. Oxygen sensing in the hypoxic response pathway: regulation of the hypoxia-inducible transcription factor. Genes Dev. 2003;17:2614–2623. - PubMed
-
- Schofield CJ, Ratcliffe PJ. Oxygen sensing by HIF hydroxylases. Nat Rev Mol Cell Biol. 2004;5:343–354. - PubMed
-
- Semenza GL. HIF-1, O(2), and the 3 PHDs: how animal cells signal hypoxia to the nucleus. Cell. 2001;107:1–3. - PubMed
-
- Ivan M, Kondo K, Yang H, Kim W, Valiando J, et al. HIFalpha targeted for VHL-mediated destruction by proline hydroxylation: implications for O2 sensing. Science. 2001;292:464–468. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
