

Similarities and differences in the structure and function of 4.1G and 4.1R135, two protein 4.1 paralogues expressed in erythroid cells
- PMID: 20812914
- PMCID: PMC4371549
- DOI: 10.1042/BJ20100041
Similarities and differences in the structure and function of 4.1G and 4.1R135, two protein 4.1 paralogues expressed in erythroid cells
Abstract
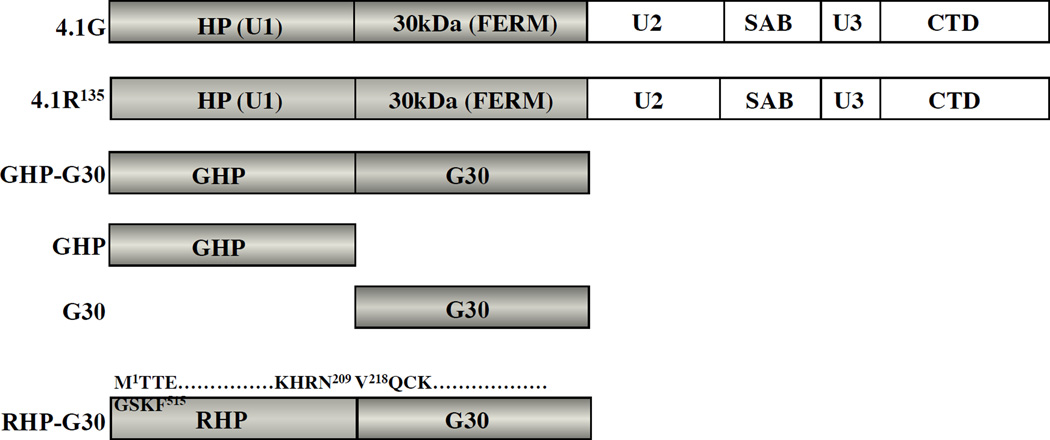
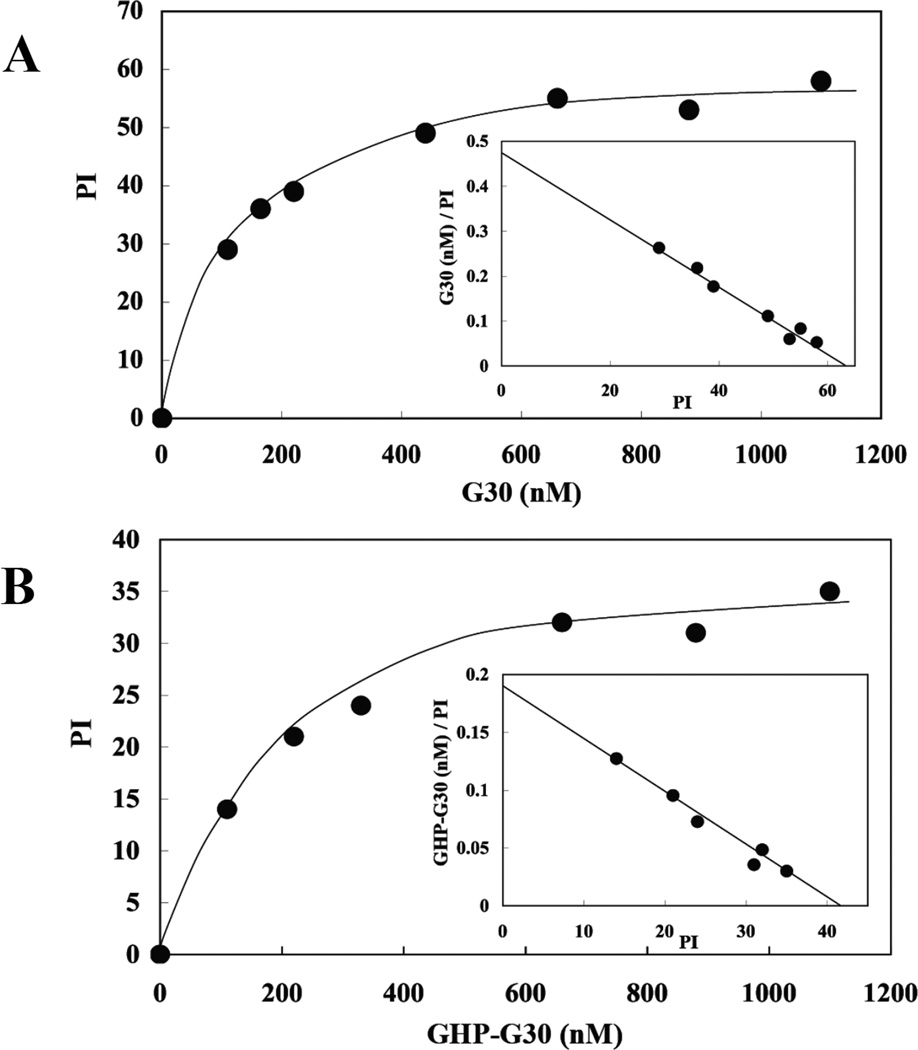
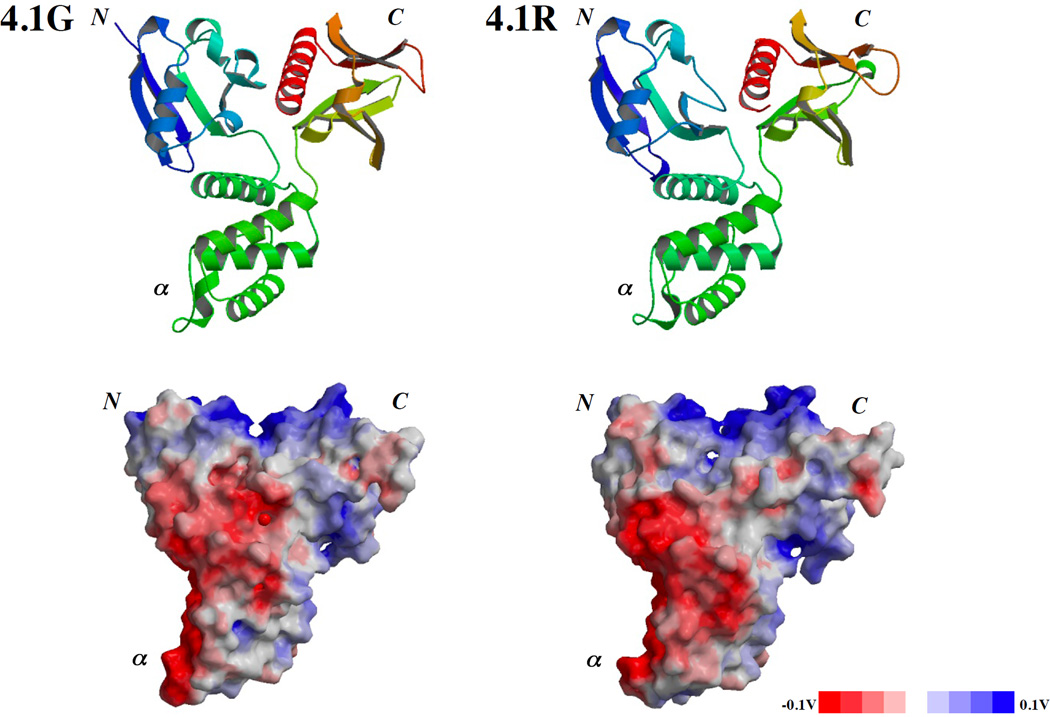
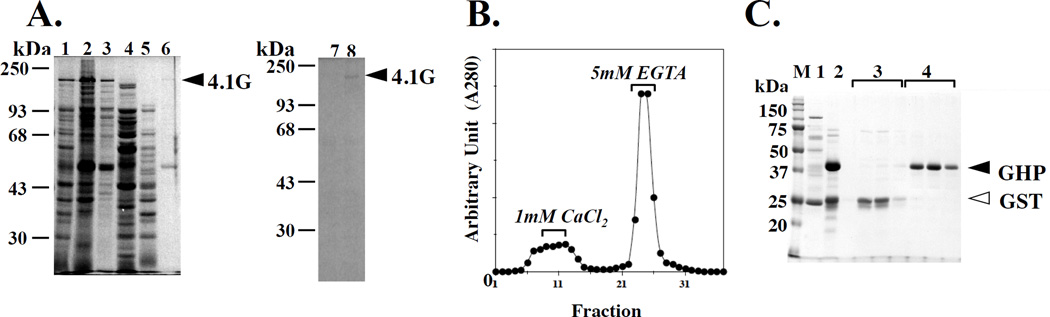
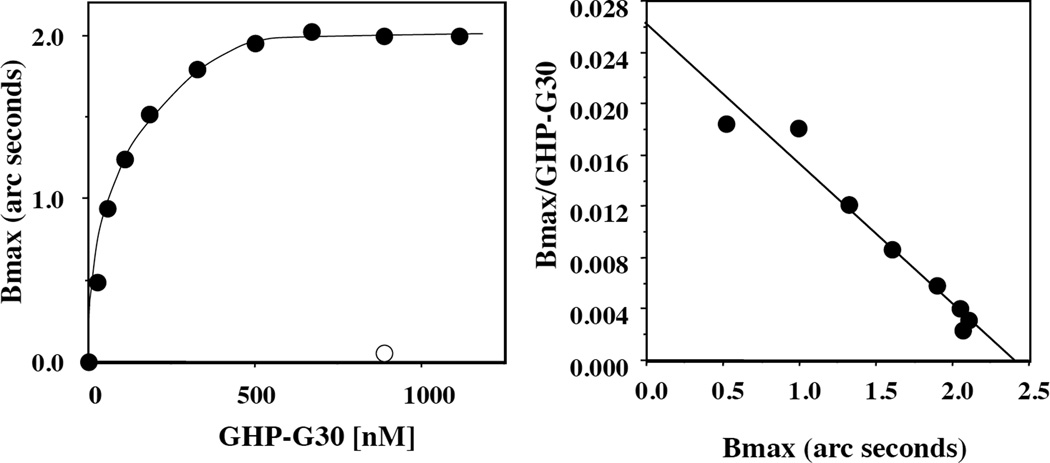
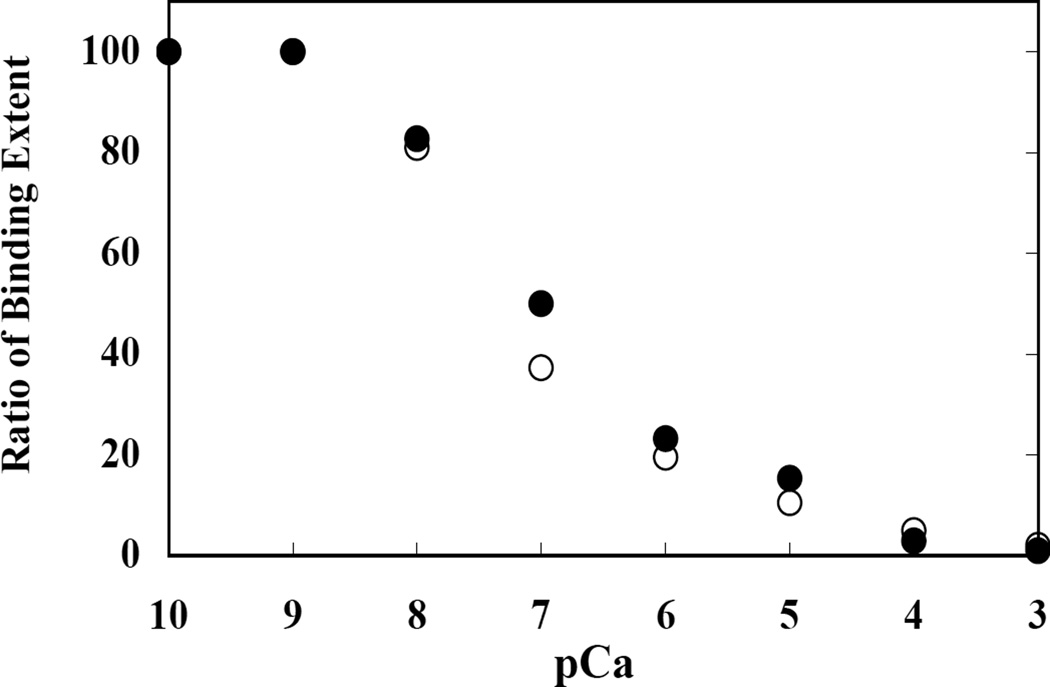
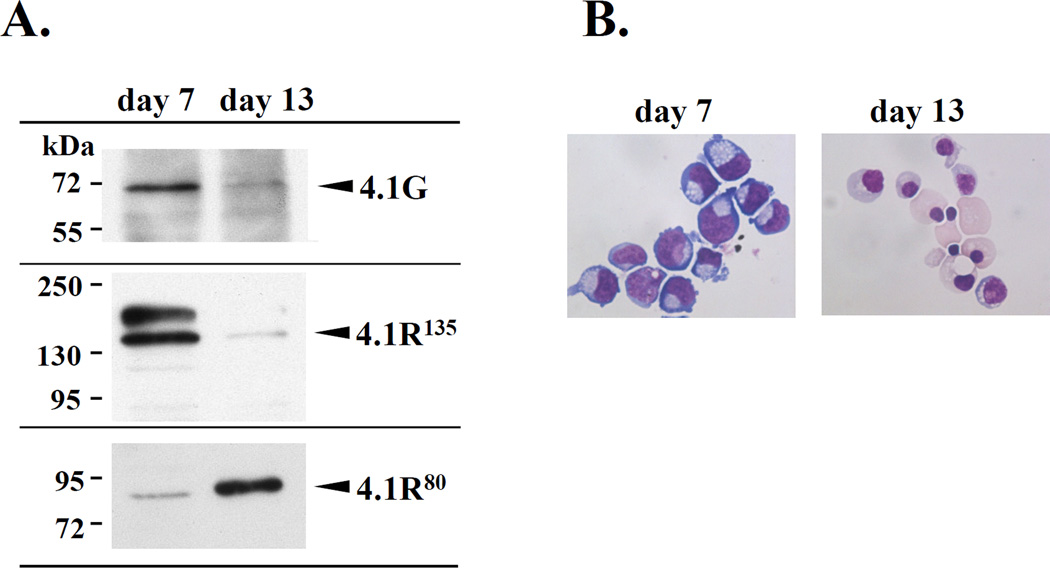
Membrane skeletal protein 4.1R is the prototypical member of a family of four highly paralogous proteins that include 4.1G, 4.1N and 4.1B. Two isoforms of 4.1R (4.1R135 and 4.1R80), as well as 4.1G, are expressed in erythroblasts during terminal differentiation, but only 4.1R80 is present in mature erythrocytes. Although the function of 4.1R isoforms in erythroid cells has been well characterized, there is little or no information on the function of 4.1G in these cells. In the present study, we performed detailed characterization of the interaction of 4.1G with various erythroid membrane proteins and the regulation of these interactions by calcium-saturated calmodulin. Like both isoforms of 4.1R, 4.1G bound to band 3, glycophorin C, CD44, p55 and calmodulin. While both 4.1G and 4.1R135 interact with similar affinity with CD44 and p55, there are significant differences in the affinity of their interaction with band 3 and glycophorin C. This difference in affinity is related to the non-conserved N-terminal headpiece region of the two proteins that is upstream of the 30 kDa membrane-binding domain that harbours the binding sites for the various membrane proteins. The headpiece region of 4.1G also contains a high-affinity calcium-dependent calmodulin-binding site that plays a key role in modulating its interaction with various membrane proteins. We suggest that expression of the two paralogues of protein 4.1 with different affinities for band 3 and glycophorin C is likely to play a role in assembly of these two membrane proteins during terminal erythroid differentiation.
Figures
References
-
- Diakowski W, Grzybek M, Sikorski AF. Protein 4.1, a component of the erythrocyte membrane skeleton and its related homologue proteins forming the protein 4.1/FERM superfamily. Folia Histochemica et Cyrobiologica. 2006;44:231–248. - PubMed
-
- Parra M, Gascard P, Walensky LD, Snyder SH, Mohandas N, Conboy JG. Cloning and characterization of 4.1G (EPB41L2), a new member of the skeletal protein 4.1 (EPB41) gene family. Genomics. 1998;49:298–306. - PubMed
-
- Parra M, Gascard P, Walensky LD, Gimm JA, Blackshaw S, Chan N, Takakuwa Y, Berger T, Lee G, Chasis JA, Snyder SH, Mohandas N, Conboy JG. Molecular and functional characterization of protein 4.1B, a novel member of the protein 4.1 family with high level, focal expression in brain. J. Biol. Chem. 2000;275:3247–3255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous