

A targeted UAS-RNAi screen in Drosophila larvae identifies wound closure genes regulating distinct cellular processes
- PMID: 20813879
- PMCID: PMC2975283
- DOI: 10.1534/genetics.110.121822
A targeted UAS-RNAi screen in Drosophila larvae identifies wound closure genes regulating distinct cellular processes
Abstract
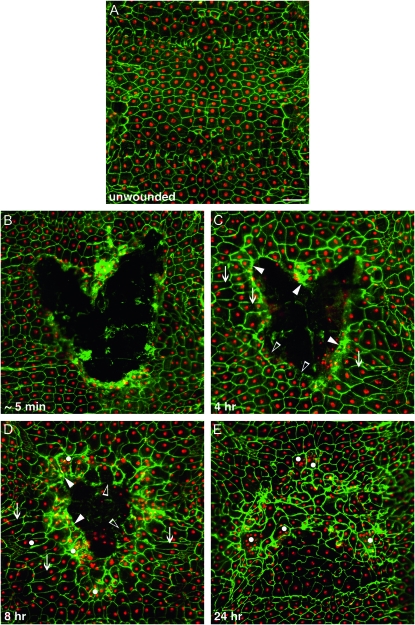
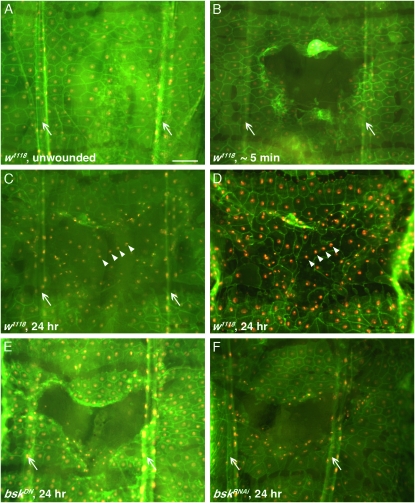
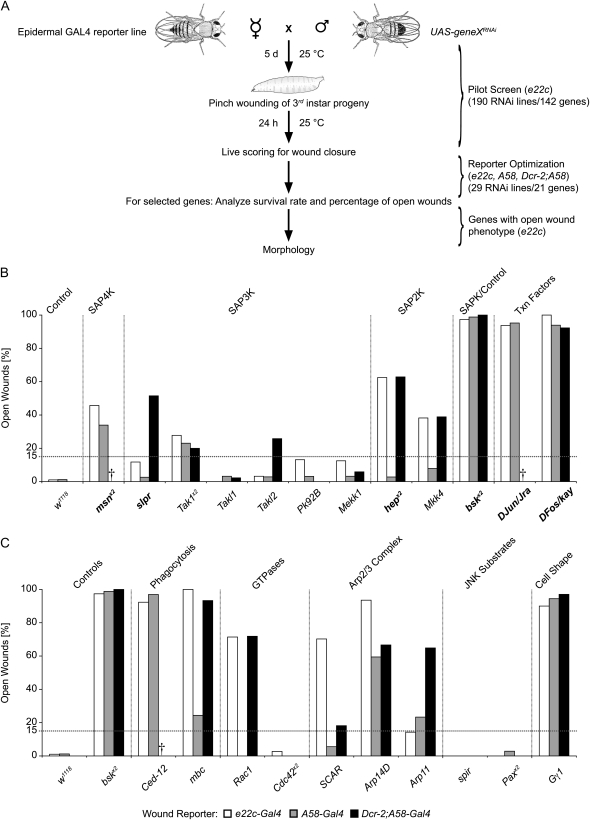
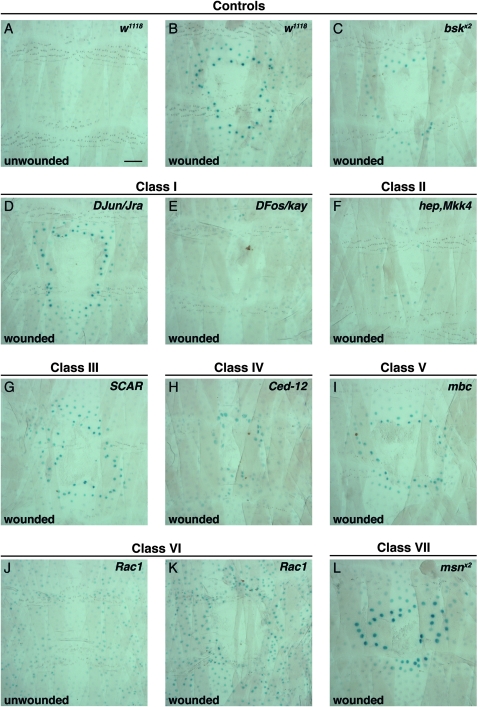
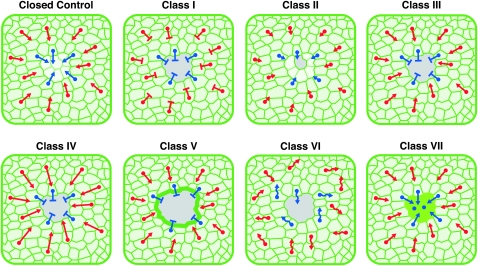
Robust mechanisms for tissue repair are critical for survival of multicellular organisms. Efficient cutaneous wound repair requires the migration of cells at the wound edge and farther back within the epidermal sheet, but the genes that control and coordinate these migrations remain obscure. This is in part because a systematic screening approach for in vivo identification and classification of postembryonic wound closure genes has yet to be developed. Here, we performed a proof-of-principle reporter-based in vivo RNAi screen in the Drosophila melanogaster larval epidermis to identify genes required for normal wound closure. Among the candidate genes tested were kinases and transcriptional mediators of the Jun N-terminal kinase (JNK) signaling pathway shown to be required for epithelial sheet migration during development. Also targeted were genes involved in actin cytoskeletal remodeling. Importantly, RNAi knockdown of both canonical and noncanonical members of the JNK pathway caused open wounds, as did several genes involved in actin cytoskeletal remodeling. Our analysis of JNK pathway components reveals redundancy among the upstream activating kinases and distinct roles for the downstream transcription factors DJun and DFos. Quantitative and qualitative morphological classification of the open wound phenotypes and evaluation of JNK activation suggest that multiple cellular processes are required in the migrating epidermal cells, including functions specific to cells at the wound edge and others specific to cells farther back within the epidermal sheet. Together, our results identify a new set of conserved wound closure genes, determine putative functional roles for these genes within the migrating epidermal sheet, and provide a template for a broader in vivo RNAi screen to discover the full complement of genes required for wound closure during larval epidermal wound healing.
Figures


Similar articles
-
cdc37 is essential for JNK pathway activation and wound closure in Drosophila.Mol Biol Cell. 2019 Oct 1;30(21):2651-2658. doi: 10.1091/mbc.E18-12-0822. Epub 2019 Sep 4. Mol Biol Cell. 2019. PMID: 31483695 Free PMC article.
-
Genetic screen in Drosophila melanogaster uncovers a novel set of genes required for embryonic epithelial repair.Genetics. 2010 Jan;184(1):129-40. doi: 10.1534/genetics.109.110288. Epub 2009 Nov 2. Genetics. 2010. PMID: 19884309 Free PMC article.
-
Yorkie regulates epidermal wound healing in Drosophila larvae independently of cell proliferation and apoptosis.Dev Biol. 2017 Jul 1;427(1):61-71. doi: 10.1016/j.ydbio.2017.05.006. Epub 2017 May 14. Dev Biol. 2017. PMID: 28514643 Free PMC article.
-
Signaling pathways directing the movement and fusion of epithelial sheets: lessons from dorsal closure in Drosophila.Differentiation. 2002 Jun;70(4-5):181-203. doi: 10.1046/j.1432-0436.2002.700408.x. Differentiation. 2002. PMID: 12147138 Review.
-
Regulating cell morphogenesis: the Drosophila Jun N-terminal kinase pathway.Genesis. 2013 Mar;51(3):147-62. doi: 10.1002/dvg.22354. Epub 2012 Nov 20. Genesis. 2013. PMID: 23109363 Review.
Cited by
-
LanB1 Cooperates With Kon-Tiki During Embryonic Muscle Migration in Drosophila.Front Cell Dev Biol. 2022 Jan 3;9:749723. doi: 10.3389/fcell.2021.749723. eCollection 2021. Front Cell Dev Biol. 2022. PMID: 35047493 Free PMC article.
-
Changes in Neuronal Signaling and Cell Stress Response Pathways are Associated with a Multigenic Response of Drosophila melanogaster to DDT Selection.Genome Biol Evol. 2017 Dec 1;9(12):3356-3372. doi: 10.1093/gbe/evx252. Genome Biol Evol. 2017. PMID: 29211847 Free PMC article.
-
Toll pathway is required for wound-induced expression of barrier repair genes in the Drosophila epidermis.Proc Natl Acad Sci U S A. 2017 Mar 28;114(13):E2682-E2688. doi: 10.1073/pnas.1613917114. Epub 2017 Mar 13. Proc Natl Acad Sci U S A. 2017. PMID: 28289197 Free PMC article.
-
Microinjection wound assay and in vivo localization of epidermal wound response reporters in Drosophila embryos.J Vis Exp. 2013 Nov 1;(81):e50750. doi: 10.3791/50750. J Vis Exp. 2013. PMID: 24300796 Free PMC article.
-
The role of transcription-independent damage signals in the initiation of epithelial wound healing.Nat Rev Mol Cell Biol. 2013 Apr;14(4):249-62. doi: 10.1038/nrm3541. Epub 2013 Feb 27. Nat Rev Mol Cell Biol. 2013. PMID: 23443750 Review.
References
-
- Baek, S. H., Y. C. Kwon, H. Lee and K. M. Choe, 2010. Rho-family small GTPases are required for cell polarization and directional sensing in Drosophila wound healing. Biochem. Biophys. Res. Commun. 394 488–492. - PubMed
-
- Bohmann, D., M. C. Ellis, L. M. Staszewski and M. Mlodzik, 1994. Drosophila Jun mediates Ras-dependent photoreceptor determination. Cell 78 973–986. - PubMed
-
- Boutros, M., H. Agaisse and N. Perrimon, 2002. Sequential activation of signaling pathways during innate immune responses in Drosophila. Dev. Cell 3 711–722. - PubMed
-
- Brand, A. H., and N. Perrimon, 1993. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118 401–415. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
