

doi: 10.1126/science.1193225.
Spiroindolones, a potent compound class for the treatment of malaria
Affiliations
- PMID: 20813948
- PMCID: PMC3050001
- DOI: 10.1126/science.1193225
Item in Clipboard
Spiroindolones, a potent compound class for the treatment of malaria
Science.
.
Abstract
Recent reports of increased tolerance to artemisinin derivatives--the most recently adopted class of antimalarials--have prompted a need for new treatments. The spirotetrahydro-beta-carbolines, or spiroindolones, are potent drugs that kill the blood stages of Plasmodium falciparum and Plasmodium vivax clinical isolates at low nanomolar concentration. Spiroindolones rapidly inhibit protein synthesis in P. falciparum, an effect that is ablated in parasites bearing nonsynonymous mutations in the gene encoding the P-type cation-transporter ATPase4 (PfATP4). The optimized spiroindolone NITD609 shows pharmacokinetic properties compatible with once-daily oral dosing and has single-dose efficacy in a rodent malaria model.
Figures
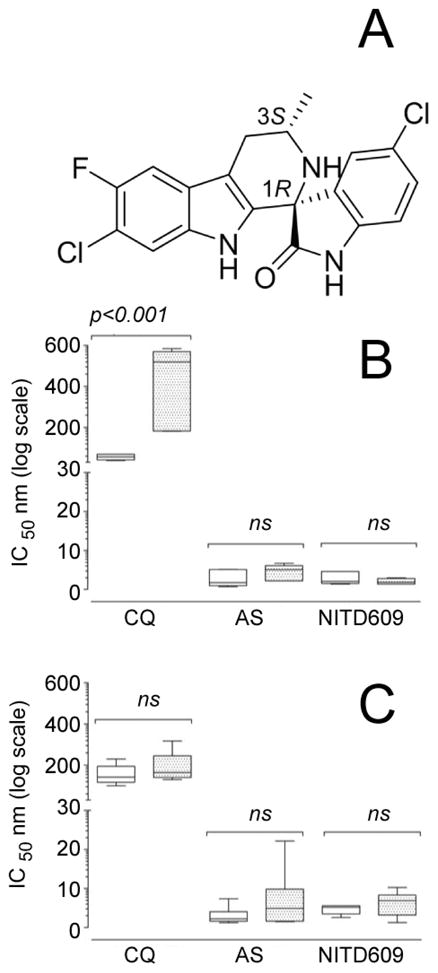
(A) Chemical structure of NITD609, showing the 1R,3S configuration that is essential for antimalarial activity. Key physicochemical properties are: solubility (pH 6.8) 39 μg/mL; logP (pH 7.4) 4.7; logD (pH 7.4) 4.6; pKa1 4.7; pKa2 10.7; polar surface area 56.92 Å2. (B) Ex vivo sensitivity of Plasmodium falciparum and (C) Plasmodium vivax (9 and 10 clinical isolates respectively) to NITD609 compared with the reference drugs chloroquine and artesunate. The antimalarial sensitivity of these two species was measured after exposing ring (unshaded boxes) and trophozoite stages (shaded boxes) to drug for 20 h. Data are shown as max–min box plots, with the solid internal line indicating median IC50 values. Inhibition of parasite growth was determined after 42 hours. Only chloroquine-treated P. vivax displayed a significant stage-specific sensitivity (p<0.001).

Spiroindolones rapidly diminish protein synthesis in the parasite. The rate of parasite protein synthesis was evaluated by monitoring [35S]-radiolabelled methionine and cysteine ([35S]-Met/Cys) incorporation into asynchronous cultures. Parasites were assayed for 1 hour in the presence of NITD609 (inverted triangle), anisomycin (diamond), cycloheximide (square), artemisinin (circle), or mefloquine (triangle), then extracted for radiographic measurements. Radiolabel incorporation was measured against inhibitor dosed over a five-log concentration range and percent incorporation was calculated by comparison to cultures assayed in the absence of inhibitor. Anisomycin and cycloheximide were included as positive controls. (A) Spiroindolone treatment rapidly diminishes protein synthesis in Dd2; however, this effect is mostly absent in (B) NITD609-RDd2 clone #2 except at very high concentration. 50% inhibition of [35S]-Met/Cys incorporation was observed with 3x and 78x IC50 of NITD609 on the NITD609-treated Dd2 wild type and NITD609-RDd2 drug-resistant clones respectively. Data are expressed in mean±SD and represent three independent experiments performed in triplicate. Similar losses of protein synthesis inhibition upon NITD609 treatment were observed in the resistant clones NITD609-RDd2 #1 and #3 respectively (see Fig. S2).
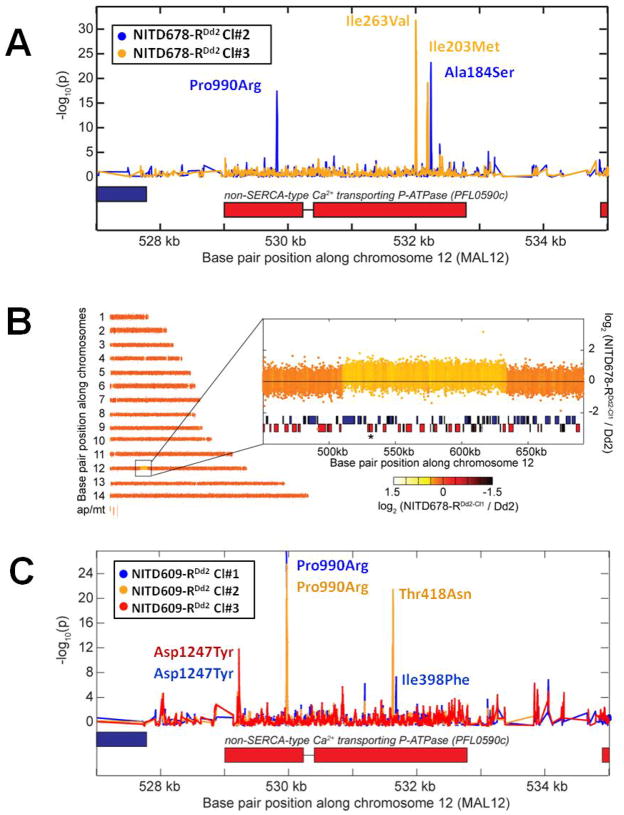
Genomic tiling arrays identified shared mutations in the pfatp4 gene (PFL0590c) in all drug-resistant parasites. (A) Distinct pairs of single nucleotide polymorphisms (SNPs) in pfatp4 were detected in NITD678-RDd2 clones #2 (blue) and #3 (orange). P-values were calculated for all probes covering pfatp4 and flanking regions; a spike in the p-value reflects a difference in hybridization between the parental clone and the drug-selected clone, a hallmark of a SNP. Direct sequencing of pfatp4 from each clone confirmed that these SNPs cause non-synonymous changes in the coding region, indicated by the red boxes. The resulting change in the primary sequence is given next to each SNP. (B) A 120kb copy number variation covering 37 genes in chromosome 12 was detected in the genome of NITD678-RDd2 clone #1. The pfatp4 gene (asterisk) was contained within this amplification. Direct sequencing of pfatp4 from this clone identified an additional non-synonymous SNP at amino acid position 223 (G223R). This mutation was continuously observed throughout numerous sequencing reads of sub-cloned pfatp4 PCR products of NITD678-RDd2 clone#1, suggesting that the mutation occurred before the amplification event and, thus, resides in all pfatp4 gene copies in the genome. (C) The three NITD609-RDd2 clones showed no evidence of copy number variants; however, each clone contained non-synonymous SNPs in pfatp4 (clone#1, blue; clone#2, orange; clone#3, red).
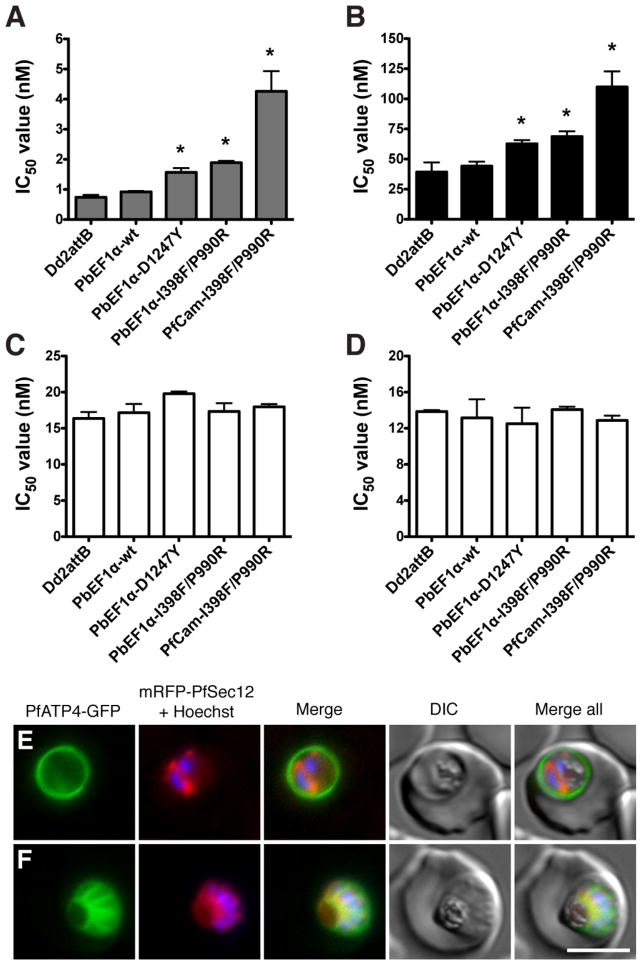
Introduction of mutant pfatp4 into Dd2attB parasites decreases susceptiblity to spiroindolones. pfatp4 transgenes harboring mutations identified in either NITD609-RDd2 clone #1 (I398F/P990R) or clone #3 (D1247Y) were individually introduced into the parental Dd2 background to evaluate the ability of the mutant protein to protect against spiroindolone activity. As a control, wild-type pfatp4 was also introduced. Expression of pfatp4 was regulated by either the P. berghei EF1α promoter (PbEF1α) or the stronger P. falciparum calmodulin promoter (PfCam). IC50 values were determined for (A) NITD609, (B) NITD678, (C) artemisinin, and (D) mefloquine. IC50 values are shown as means±stdev and were derived from three independent experiments performed in quadruplicate with the SYBR Green-based cell proliferation assay (12). Statistical significance was calculated using a two-tailed unpaired t test, comparing transgenic pfatp4 lines to the Dd2attB parental line: *p<0.0001. (E, F) Localization of PfATP4 to the parasite plasma membrane. Transgenic parasites co-expressing PfATP4-GFP and an ER marker, mRFP-PfSec12 were labeled with Hoechst 33382 to visualize the nucleus. PfATP4-GFP was observed at the parasite plasma membrane in (E) early schizont (two nuclei) and (F) late-segmented schizont parasites. Bar = 5 μm.
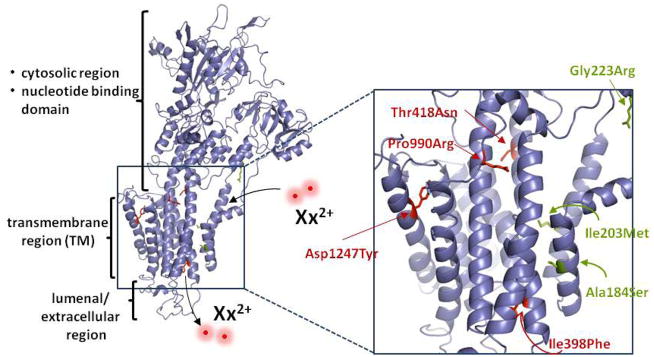
Resistance-associated SNPs map to the predicted transmembrane region of PfATP4. A homology model of PfATP4 was generated in SWISS-MODEL based on the crystal structure of the rabbit SERCA pump. Amino acid alignment analysis by EMBOSS (41) revealed 30% identity and 48% similarity between these proteins. Residues corresponding to resistance-associated mutations are indicated in red for NITD609-RDd2 and in green for NITD678-RDd2. These mutations mapped to the putative transmembrane helices. The sites of divalent cation entry and exit are indicated as Xx2+.
Comment in
-
Microbiology. Is the tide turning for new malaria medicines?Science. 2010 Sep 3;329(5996):1153-4. doi: 10.1126/science.1194923. Science. 2010. PMID: 20813940 No abstract available.
-
Antimalarial drugs: Speeding to a new lead.Nat Rev Drug Discov. 2010 Nov;9(11):842. doi: 10.1038/nrd3301. Nat Rev Drug Discov. 2010. PMID: 21031000 No abstract available.
References
-
- WHO. World Malaria Report. 2009. http://www.who.int/malaria/world_malaria_report_2009/en/
-
- Noedl H, et al. N Engl J Med. 2008;359:2619. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Chemical Information
Medical
Molecular Biology Databases
