

Coupling between microtubule sliding, plus-end growth and spindle length revealed by kinesin-8 depletion
- PMID: 20814910
- PMCID: PMC2998535
- DOI: 10.1002/cm.20482
Coupling between microtubule sliding, plus-end growth and spindle length revealed by kinesin-8 depletion
Abstract
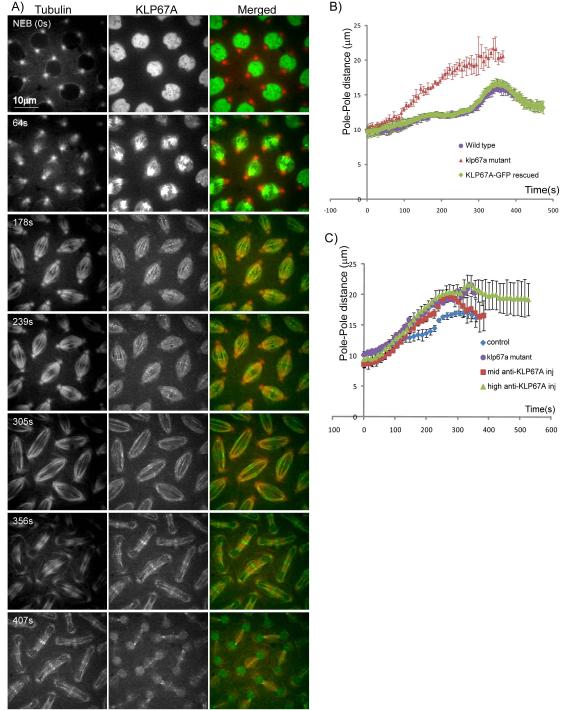
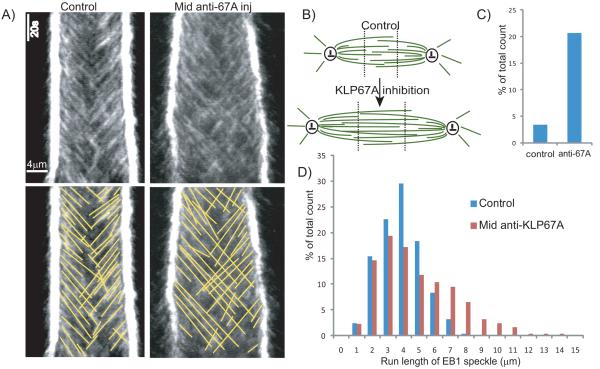
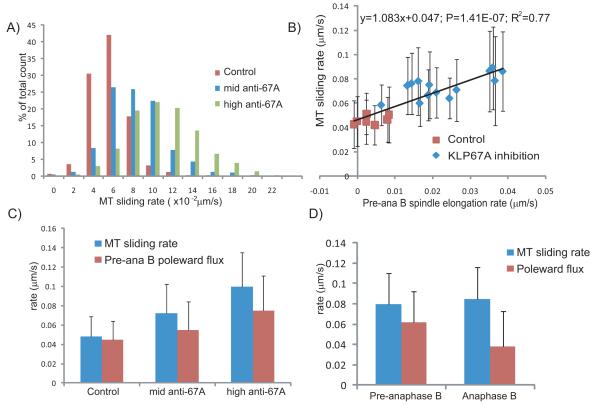
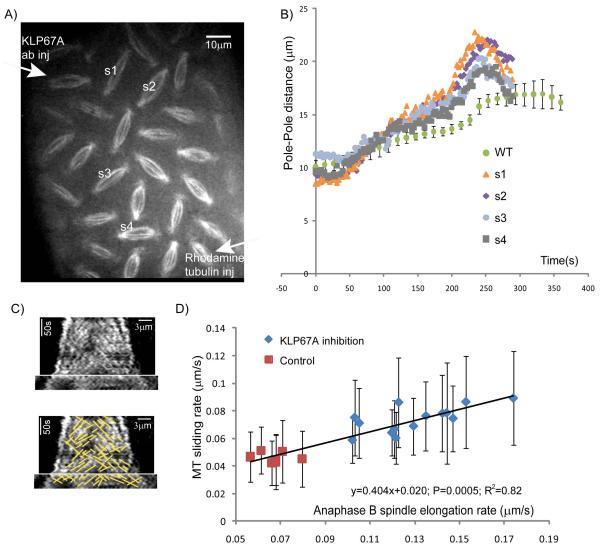
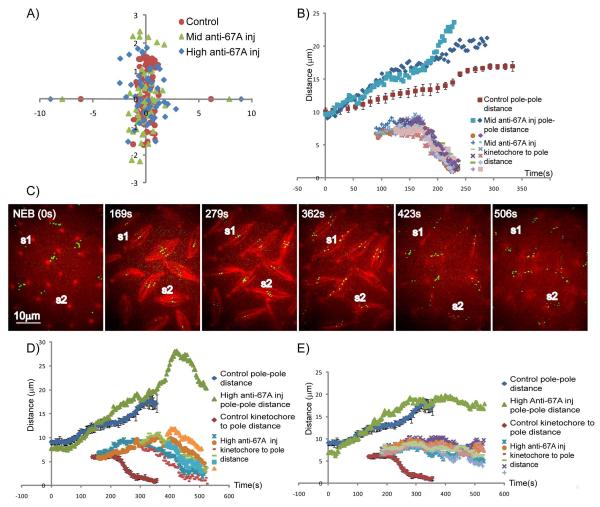
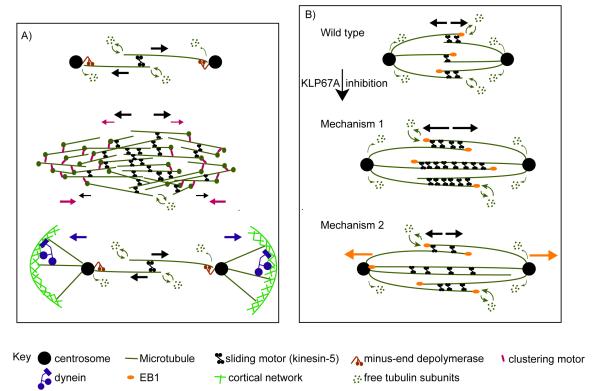
Mitotic spindle length control requires coordination between microtubule (MT) dynamics and motor-generated forces. To investigate how MT plus-end polymerization contributes to spindle length in Drosophila embryos, we studied the dynamics of the MT plus-end depolymerase, kinesin-8, and the effects of kinesin-8 inhibition using mutants and antibody microinjection. As expected, kinesin-8 was found to contribute to anaphase A. Furthermore, kinesin-8 depletion caused: (i) excessive polymerization of interpolar (ip) MT plus ends, which "overgrow" to penetrate distal half spindles; (ii) an increase in the poleward ipMT sliding rate that is coupled to MT plus-end polymerization; (iii) premature spindle elongation during metaphase/anaphase A; and (iv) an increase in the anaphase B spindle elongation rate which correlates linearly with the MT sliding rate. This is best explained by a revised "ipMT sliding/minus-end depolymerization" model for spindle length control which incorporates a coupling between ipMT plus end dynamics and the outward ipMT sliding that drives poleward flux and spindle elongation.
© 2010 Wiley-Liss, Inc.
Figures
Similar articles
-
Patronin mediates a switch from kinesin-13-dependent poleward flux to anaphase B spindle elongation.J Cell Biol. 2013 Oct 14;203(1):35-46. doi: 10.1083/jcb.201306001. Epub 2013 Oct 7. J Cell Biol. 2013. PMID: 24100293 Free PMC article.
-
Mitotic motors and chromosome segregation: the mechanism of anaphase B.Biochem Soc Trans. 2011 Oct;39(5):1149-53. doi: 10.1042/BST0391149. Biochem Soc Trans. 2011. PMID: 21936780 Review.
-
Kinesin-5-dependent poleward flux and spindle length control in Drosophila embryo mitosis.Mol Biol Cell. 2009 Mar;20(6):1749-62. doi: 10.1091/mbc.e08-10-1033. Epub 2009 Jan 21. Mol Biol Cell. 2009. PMID: 19158379 Free PMC article.
-
Mechanism for Anaphase B: Evaluation of "Slide-and-Cluster" versus "Slide-and-Flux-or-Elongate" Models.Biophys J. 2015 Apr 21;108(8):2007-18. doi: 10.1016/j.bpj.2015.03.018. Biophys J. 2015. PMID: 25902440 Free PMC article.
-
Anaphase B.Biology (Basel). 2016 Dec 8;5(4):51. doi: 10.3390/biology5040051. Biology (Basel). 2016. PMID: 27941648 Free PMC article. Review.
Cited by
-
Patronin mediates a switch from kinesin-13-dependent poleward flux to anaphase B spindle elongation.J Cell Biol. 2013 Oct 14;203(1):35-46. doi: 10.1083/jcb.201306001. Epub 2013 Oct 7. J Cell Biol. 2013. PMID: 24100293 Free PMC article.
-
The Drosophila orthologue of the INT6 onco-protein regulates mitotic microtubule growth and kinetochore structure.PLoS Genet. 2017 May 15;13(5):e1006784. doi: 10.1371/journal.pgen.1006784. eCollection 2017 May. PLoS Genet. 2017. PMID: 28505193 Free PMC article.
-
Aurora B and Kif2A control microtubule length for assembly of a functional central spindle during anaphase.J Cell Biol. 2013 Aug 19;202(4):623-36. doi: 10.1083/jcb.201302123. J Cell Biol. 2013. PMID: 23960144 Free PMC article.
-
Microtubule rescue at midzone edges promotes overlap stability and prevents spindle collapse during anaphase B.Elife. 2022 Mar 16;11:e72630. doi: 10.7554/eLife.72630. Elife. 2022. PMID: 35293864 Free PMC article.
-
The emergence of sarcomeric, graded-polarity and spindle-like patterns in bundles of short cytoskeletal polymers and two opposite molecular motors.J Phys Condens Matter. 2011 Sep 21;23(37):374102. doi: 10.1088/0953-8984/23/37/374102. Epub 2011 Aug 23. J Phys Condens Matter. 2011. PMID: 21862843 Free PMC article.
References
-
- Brust-Mascher I, Scholey JM. Microinjection Techniques for Studying Mitosis in the Drosophila melanogaster Syncytial Embryo. JoVE. 2009;31 http://www.jove.com/index/details.stp?id=1382, DOI: 10.3791/1382. - PMC - PubMed
-
- Burbank KS, Mitchison TJ, Fisher DS. Slide-and-cluster models for spindle assembly. Curr Biol. 2007;17:1373–1383. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
