

Female sexual arousal in amphibians
- PMID: 20816968
- PMCID: PMC3008597
- DOI: 10.1016/j.yhbeh.2010.08.015
Female sexual arousal in amphibians
Abstract
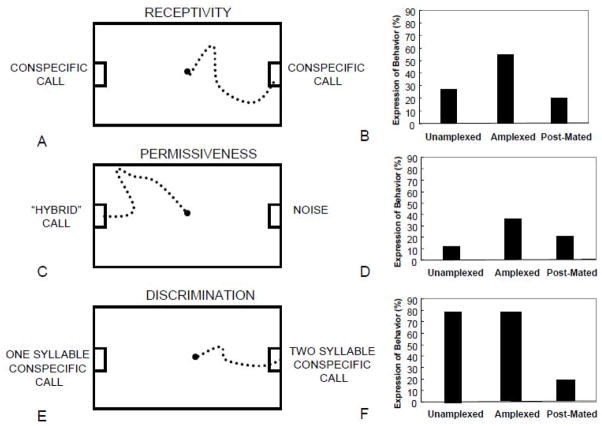
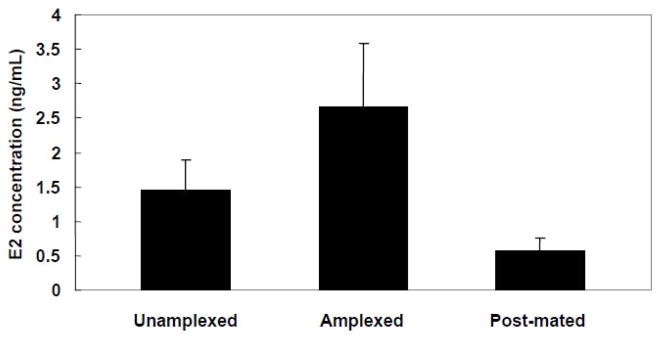
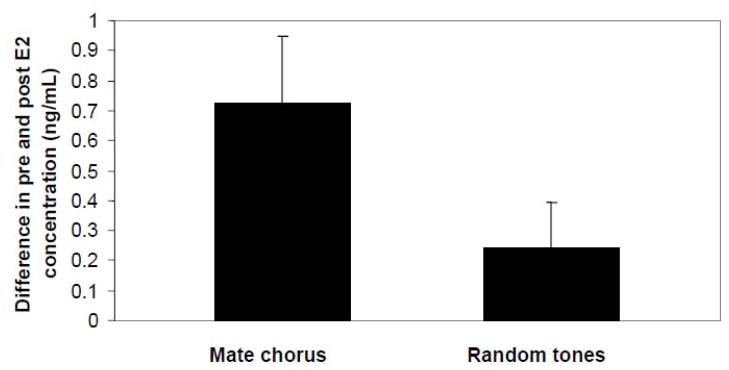
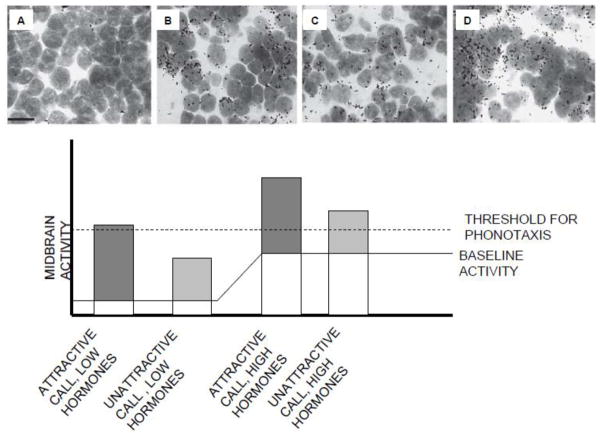
Rather than being a static, species specific trait, reproductive behavior in female amphibians is variable within an individual during the breeding season when females are capable of reproductive activity. Changes in receptivity coincide with changes in circulating estrogen. Estrogen is highest at the point when females are ready to choose a male and lay eggs. At this time female receptivity (her probability of responding to a male vocal signal) is highest and her selectivity among conspecific calls (measured by her probability of responding to a degraded or otherwise usually unattractive male signal) is lowest. These changes occur even though females retain the ability to discriminate different acoustic characteristics of various conspecific calls. After releasing her eggs, female amphibians quickly become less receptive and more choosy in terms of their responses to male sexual advertisement signals. Male vocal signals stimulate both behavior and estrogen changes in amphibian females making mating more probable. The changes in female reproductive behavior are the same as those generally accepted as indicative of a change in female sexual arousal leading to copulation. They are situationally triggered, gated by interactions with males, and decline with the consummation of sexual reproduction with a chosen male. The changes can be triggered by either internal physiological state or by the presence of stimuli presented by males, and the same stimuli change both behavior and physiological (endocrine) state in such a way as to make acceptance of a male more likely. Thus amphibian females demonstrate many of the same general characteristics of changing female sexual state that in mammals indicate sexual arousal.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Historical perspective: Hormonal regulation of behaviors in amphibians.Horm Behav. 2005 Nov;48(4):373-83. doi: 10.1016/j.yhbeh.2005.05.011. Epub 2005 Jun 29. Horm Behav. 2005. PMID: 15992801 Review.
-
Sexual hearing: the influence of sex hormones on acoustic communication in frogs.Hear Res. 2009 Jun;252(1-2):15-20. doi: 10.1016/j.heares.2009.01.001. Epub 2009 Jan 17. Hear Res. 2009. PMID: 19272318 Free PMC article. Review.
-
Who rises to the challenge? Testing the Challenge Hypothesis in fish, amphibians, reptiles, and mammals.Horm Behav. 2020 Jul;123:104537. doi: 10.1016/j.yhbeh.2019.06.001. Epub 2019 Jun 22. Horm Behav. 2020. PMID: 31181193 Review.
-
Mating vocalizations of female frogs: control and evolutionary mechanisms.Brain Behav Evol. 1999;53(4):187-97. doi: 10.1159/000006594. Brain Behav Evol. 1999. PMID: 10343085 Review.
-
Female reproductive state influences the auditory midbrain response.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2009 Apr;195(4):341-9. doi: 10.1007/s00359-008-0410-7. Epub 2009 Jan 30. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2009. PMID: 19184041
Cited by
-
Electroencephalographic signals synchronize with behaviors and are sexually dimorphic during the light-dark cycle in reproductive frogs.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2014 Feb;200(2):117-27. doi: 10.1007/s00359-013-0866-y. Epub 2013 Dec 12. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2014. PMID: 24337372
-
A cretaceous frog with eggs from northwestern China provides fossil evidence for sexual maturity preceding skeletal maturity in anurans.Proc Biol Sci. 2024 Feb 14;291(2016):20232320. doi: 10.1098/rspb.2023.2320. Epub 2024 Feb 7. Proc Biol Sci. 2024. PMID: 38320608 Free PMC article.
-
We Made Your Bed, Why Won't You Lie in It? Food Availability and Disease May Affect Reproductive Output of Reintroduced Frogs.PLoS One. 2016 Jul 27;11(7):e0159143. doi: 10.1371/journal.pone.0159143. eCollection 2016. PLoS One. 2016. PMID: 27463095 Free PMC article.
-
Reproductive behavior drives female space use in a sedentary Neotropical frog.PeerJ. 2020 Apr 17;8:e8920. doi: 10.7717/peerj.8920. eCollection 2020. PeerJ. 2020. PMID: 32337103 Free PMC article.
-
Socially modulated cell proliferation is independent of gonadal steroid hormones in the brain of the adult green treefrog (Hyla cinerea).Brain Behav Evol. 2012;79(3):170-80. doi: 10.1159/000335037. Epub 2012 Jan 20. Brain Behav Evol. 2012. PMID: 22269468 Free PMC article.
References
-
- Beach FA. Sexual attractivity, proceptivity, and receptivity in female mammals. Horm Behav. 1976;7:105–138. - PubMed
-
- Bosch J, Boyero L. Reproductive stage and phonotatic preferences of female midwife toads (Alytes cisternasii) Behav Ecol Sociobiol. 2004;55:251–256.
-
- Boyd SK. Sexual differences in hormonal control of release calls in bullfrogs. Horm Behav. 1992;26:522–535. - PubMed
-
- Boyd SK. Arginine vasotocin facilitation of advertisement calling and call phonotaxis in bullfrogs. Horm Behav. 1994;28:232–240. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
