

Iron-export ferroxidase activity of β-amyloid precursor protein is inhibited by zinc in Alzheimer's disease
- PMID: 20817278
- PMCID: PMC2943017
- DOI: 10.1016/j.cell.2010.08.014
Iron-export ferroxidase activity of β-amyloid precursor protein is inhibited by zinc in Alzheimer's disease
Abstract
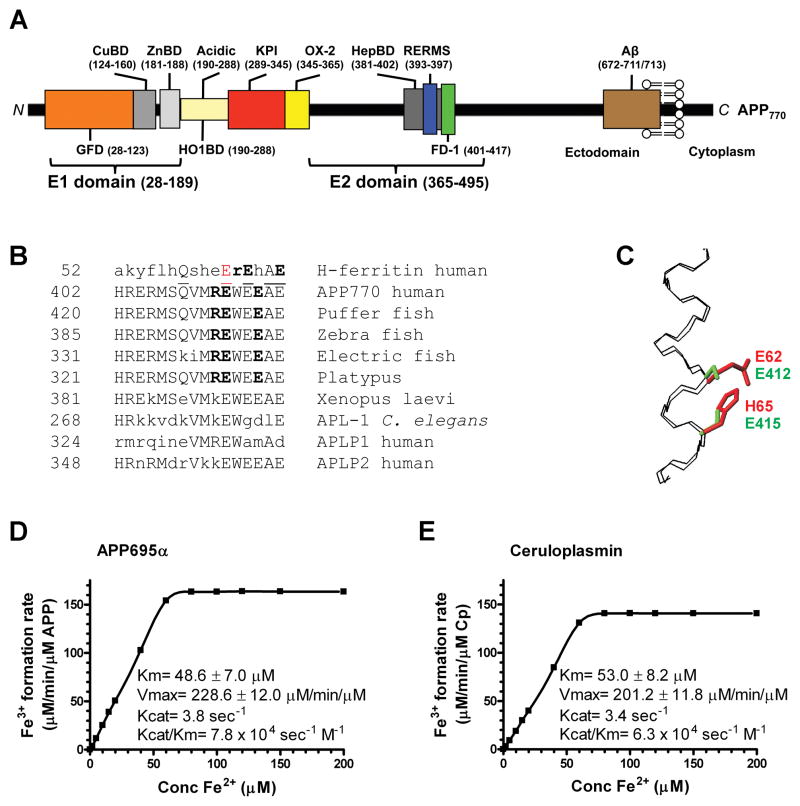
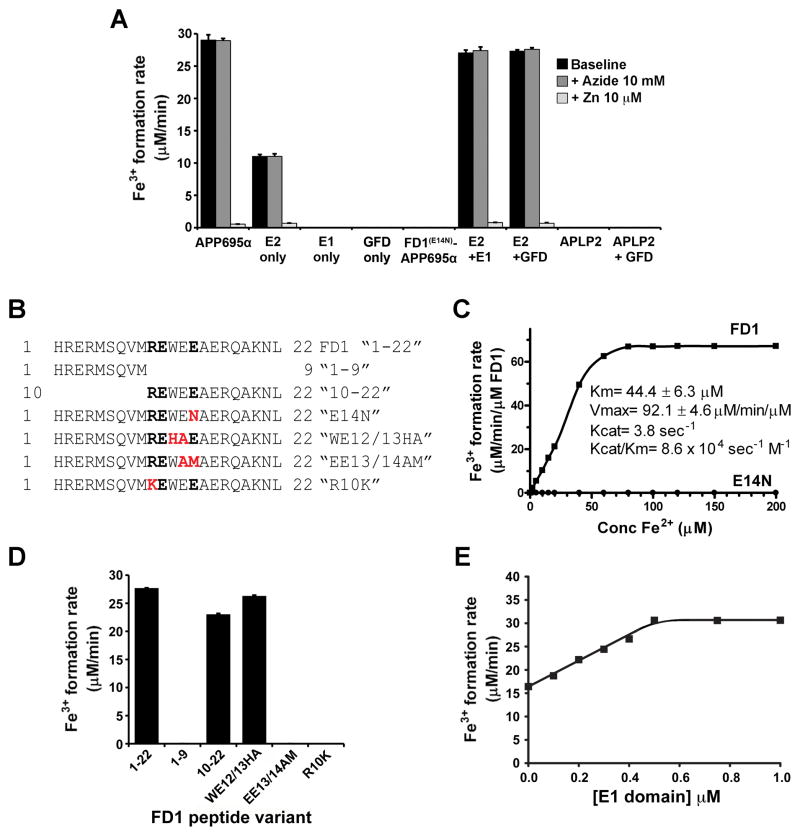
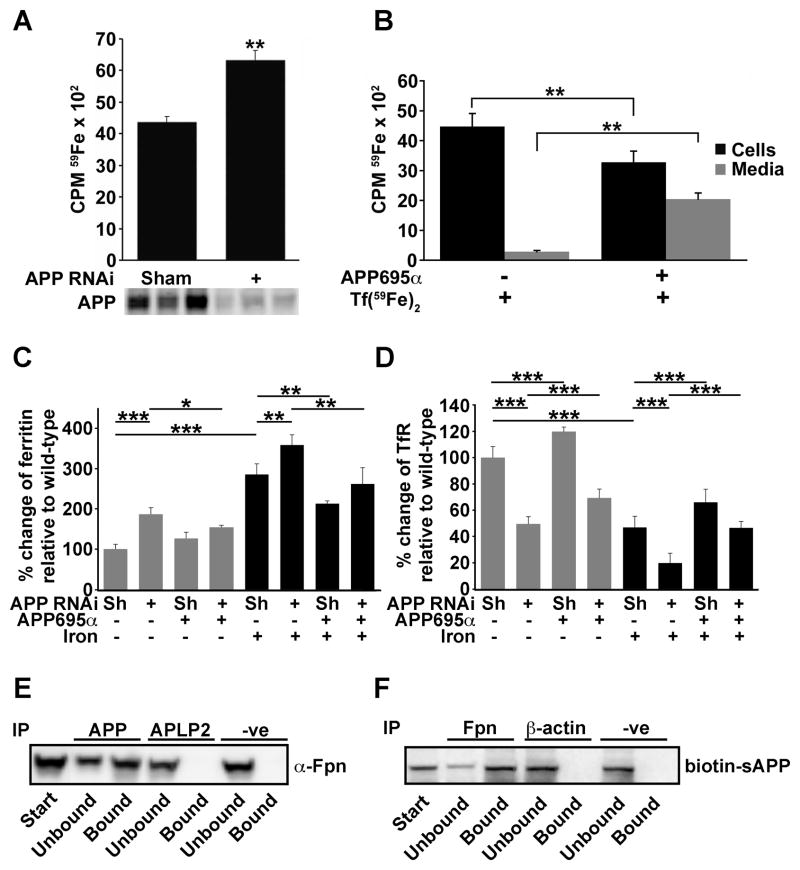
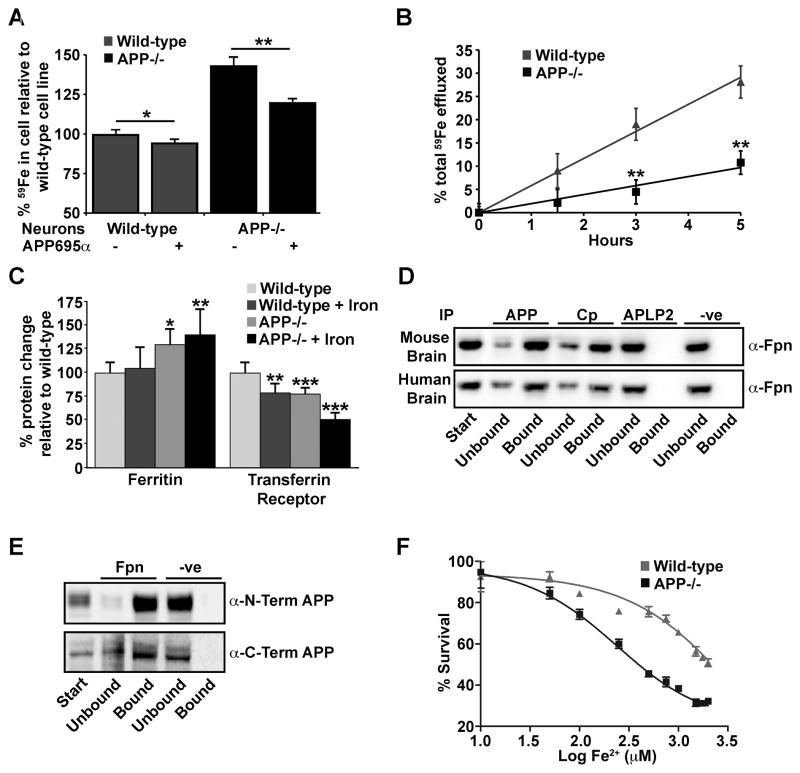
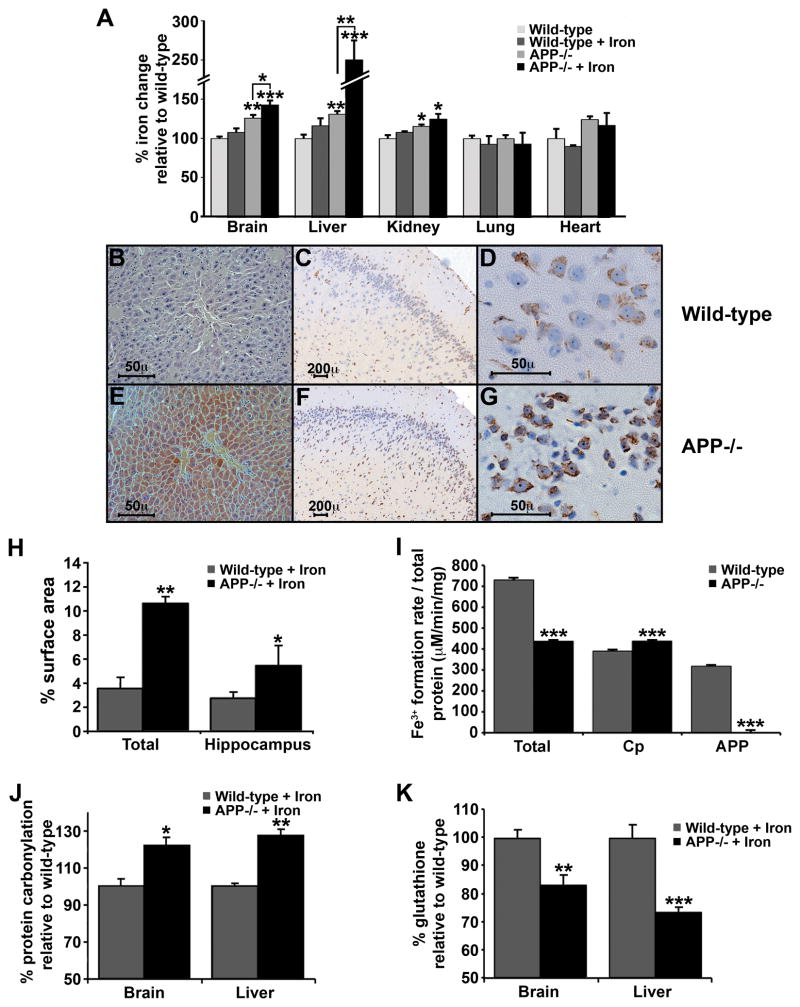
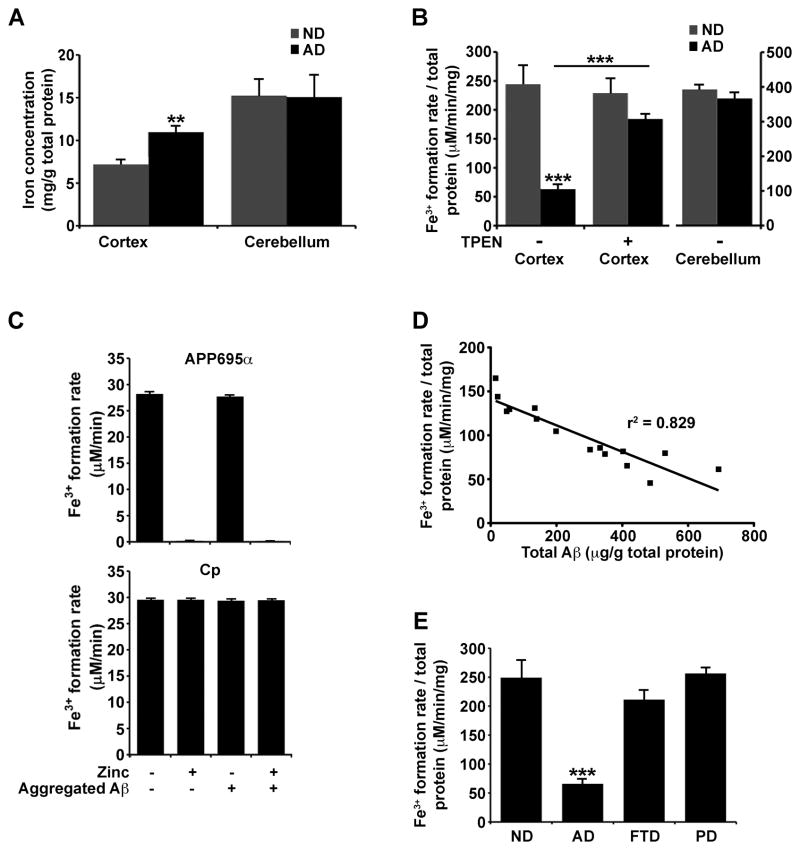
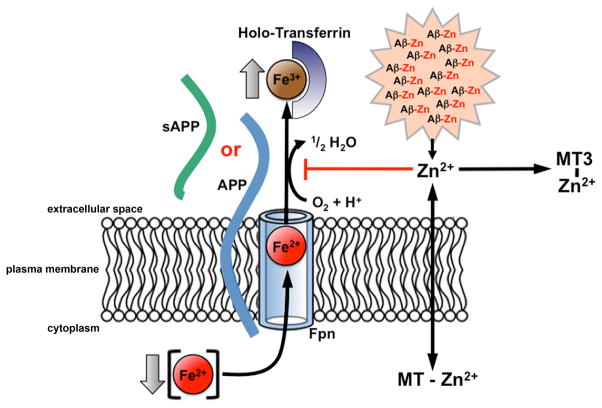
Alzheimer's Disease (AD) is complicated by pro-oxidant intraneuronal Fe(2+) elevation as well as extracellular Zn(2+) accumulation within amyloid plaque. We found that the AD β-amyloid protein precursor (APP) possesses ferroxidase activity mediated by a conserved H-ferritin-like active site, which is inhibited specifically by Zn(2+). Like ceruloplasmin, APP catalytically oxidizes Fe(2+), loads Fe(3+) into transferrin, and has a major interaction with ferroportin in HEK293T cells (that lack ceruloplasmin) and in human cortical tissue. Ablation of APP in HEK293T cells and primary neurons induces marked iron retention, whereas increasing APP695 promotes iron export. Unlike normal mice, APP(-/-) mice are vulnerable to dietary iron exposure, which causes Fe(2+) accumulation and oxidative stress in cortical neurons. Paralleling iron accumulation, APP ferroxidase activity in AD postmortem neocortex is inhibited by endogenous Zn(2+), which we demonstrate can originate from Zn(2+)-laden amyloid aggregates and correlates with Aβ burden. Abnormal exchange of cortical zinc may link amyloid pathology with neuronal iron accumulation in AD.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Adlard PA, Cherny RA, Finkelstein DI, Gautier E, Robb E, Cortes M, Volitakis I, Liu X, Smith JP, Perez K, et al. Rapid restoration of cognition in Alzheimer’s transgenic mice with 8-hydroxy quinoline analogs is associated with decreased interstitial Abeta. Neuron. 2008;59:43–55. - PubMed
-
- Bakker GR, Boyer RF. Iron incorporation into apoferritin. The role of apoferritin as a ferroxidase. J Biol Chem. 1986;261:13182–13185. - PubMed
-
- Bartzokis G, Sultzer D, Mintz J, Holt LE, Marx P, Phelan CK, Marder SR. In vivo evaluation of brain iron in Alzheimer’s disease and normal subjects using MRI. Biol Psychiatry. 1994b;35:480–487. - PubMed
-
- Bartzokis G, Tishler TA. MRI evaluation of basal ganglia ferritin iron and neurotoxicity in Alzheimer’s and Huntingon’s disease. Cell Mol Biol (Noisy-le-grand) 2000;46:821–833. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
