

Diverse evolutionary paths to cell adhesion
- PMID: 20817460
- PMCID: PMC2991404
- DOI: 10.1016/j.tcb.2010.08.002
Diverse evolutionary paths to cell adhesion
Abstract
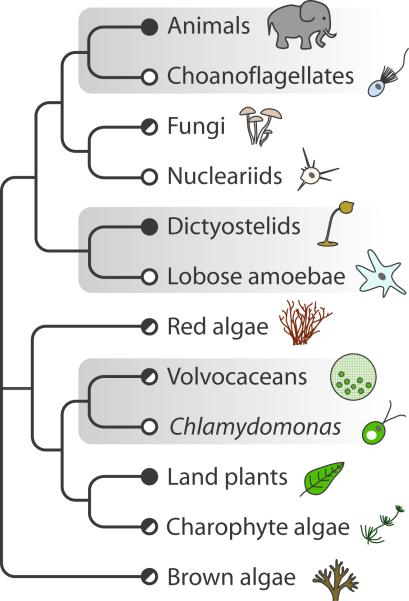
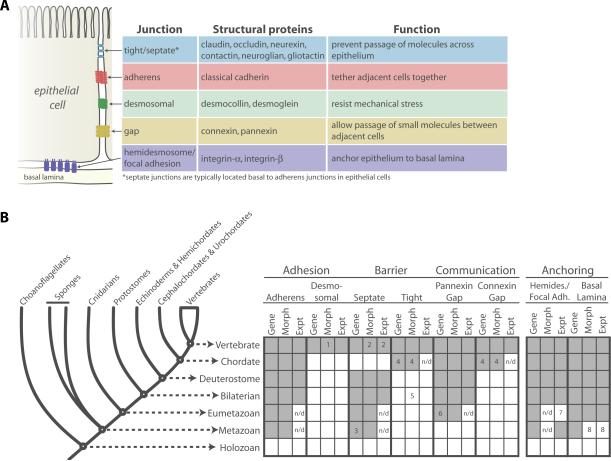
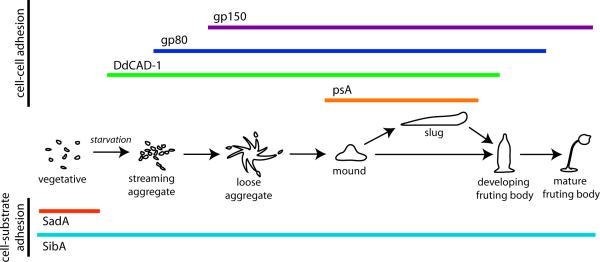
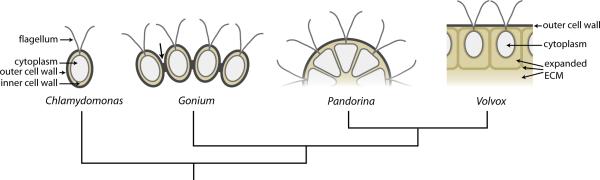
The morphological diversity of animals, fungi, plants, and other multicellular organisms stems from the fact that each lineage acquired multicellularity independently. A prerequisite for each origin of multicellularity was the evolution of mechanisms for stable cell-cell adhesion or attachment. Recent advances in comparative genomics and phylogenetics provide critical insights into the evolutionary foundations of cell adhesion. Reconstructing the evolution of cell junction proteins in animals and their unicellular relatives exemplifies the roles of co-option and innovation. Comparative studies of volvocine algae reveal specific molecular changes that accompanied the evolution of multicellularity in Volvox. Comparisons between animals and Dictyostelium show how commonalities and differences in the biology of unicellular ancestors influenced the evolution of adhesive mechanisms. Understanding the unicellular ancestry of cell adhesion helps illuminate the basic cell biology of multicellular development in modern organisms.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
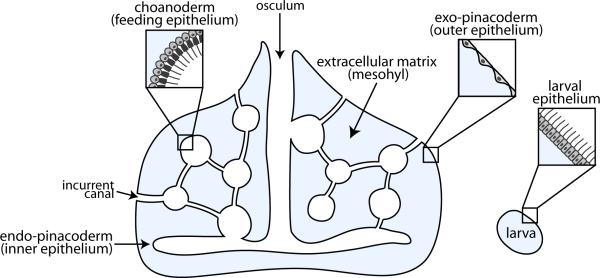
References
-
- Jarvis MC, et al. Intercellular adhesion and cell separation in plants. Plant Cell Environ. 2003;26:977–989.
-
- Gumbiner BM. Cell adhesion: the molecular basis of tissue architecture and morphogenesis. Cell. 1996;84:345–357. - PubMed
-
- King N. The unicellular ancestry of animal development. Dev. Cell. 2004;7:313–325. - PubMed
-
- Baldauf SL. The deep roots of eukaryotes. Science. 2003;300:1703–1706. - PubMed
-
- Carroll SB. Chance and necessity: the evolution of morphological complexity and diversity. Nature. 2001;409:1102–1109. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
