

Molecular basis of LMAN1 in coordinating LMAN1-MCFD2 cargo receptor formation and ER-to-Golgi transport of FV/FVIII
- PMID: 20817851
- PMCID: PMC3031414
- DOI: 10.1182/blood-2010-04-278325
Molecular basis of LMAN1 in coordinating LMAN1-MCFD2 cargo receptor formation and ER-to-Golgi transport of FV/FVIII
Abstract
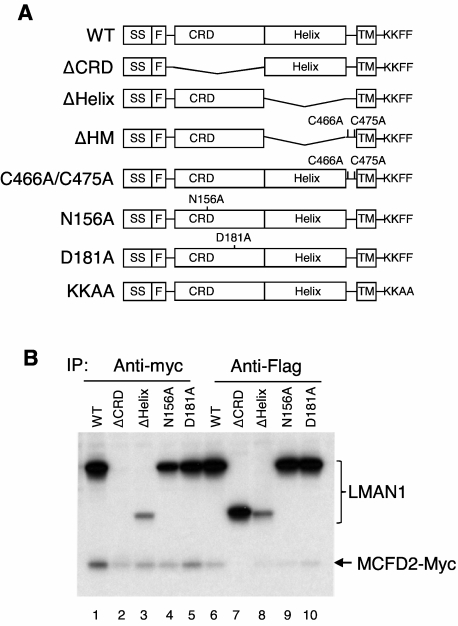
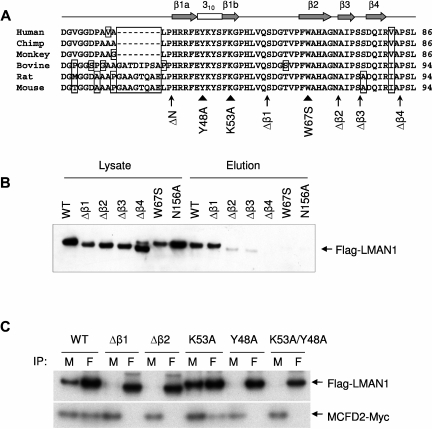
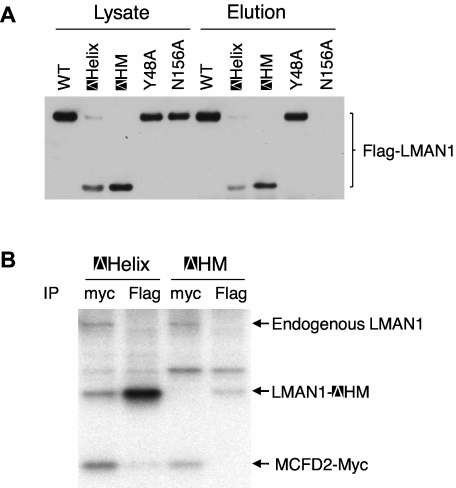
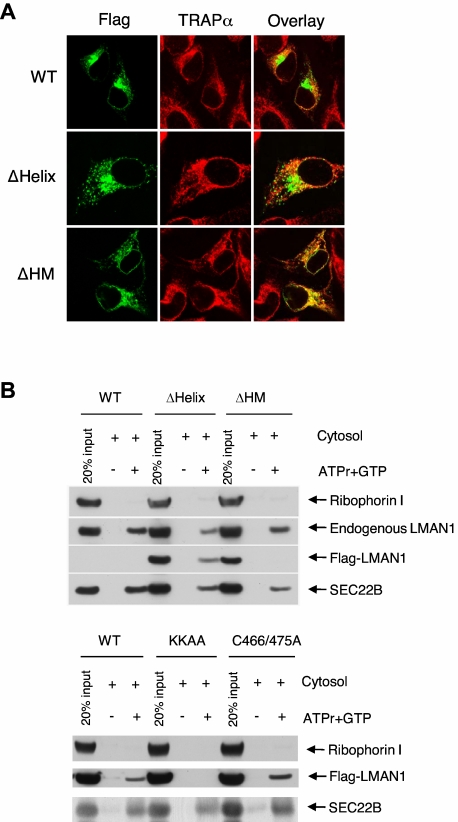
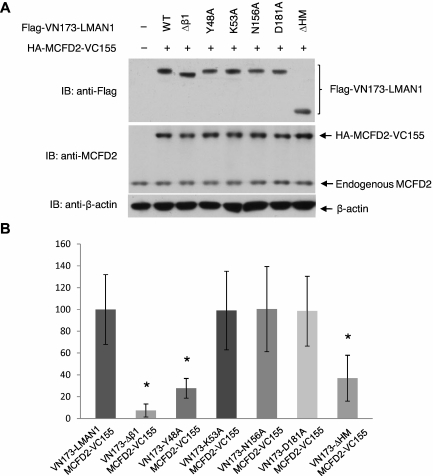
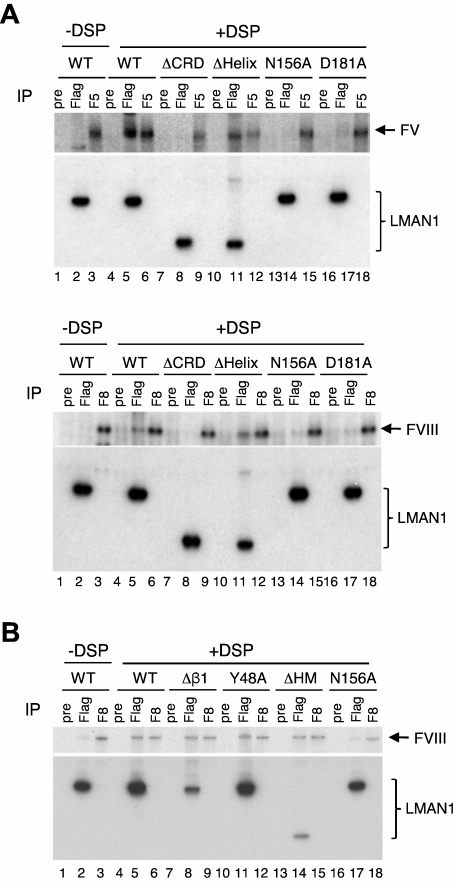
The LMAN1-MCFD2 (lectin, mannose binding 1/multiple coagulation factor deficiency protein 2) cargo receptor complex transports coagulation factors V (FV) and VIII (FVIII) from the endoplasmic reticulum (ER) to the ER-Golgi intermediate compartment (ERGIC). LMAN1 (ERGIC-53) is a hexameric transmembrane protein with a carbohydrate recognition domain (CRD) on the ER luminal side. Here, we show that mutations in the first beta sheet of the CRD abolish MCFD2 binding without affecting the mannose binding, suggesting that LMAN1 interacts with MCFD2 through its N-terminal beta sheet, consistent with recently reported crystal structures of the CRD-MCFD2 complex. Mutations in the Ca(2+)- and sugar-binding sites of the CRD disrupt FV and FVIII interactions, without affecting MCFD2 binding. This interaction is independent of MCFD2, as LMAN1 mutants defective in MCFD2 binding can still interact with FVIII. Thus, the CRD of LMAN1 contains distinct, separable binding sites for both its partner protein (MCFD2) and the cargo proteins (FV/FVIII). Monomeric LMAN1 mutants are defective in ER exit and unable to interact with MCFD2, suggesting that the oligomerization of LMAN1 is necessary for its cargo receptor function. These results point to a central role of LMAN1 in regulating the binding in the ER and the subsequent release in the ERGIC of FV and FVIII.
Figures
Similar articles
-
Structural characterization of carbohydrate binding by LMAN1 protein provides new insight into the endoplasmic reticulum export of factors V (FV) and VIII (FVIII).J Biol Chem. 2013 Jul 12;288(28):20499-509. doi: 10.1074/jbc.M113.461434. Epub 2013 May 24. J Biol Chem. 2013. PMID: 23709226 Free PMC article.
-
EF-hand domains of MCFD2 mediate interactions with both LMAN1 and coagulation factor V or VIII.Blood. 2010 Feb 4;115(5):1081-7. doi: 10.1182/blood-2009-09-241877. Epub 2009 Dec 9. Blood. 2010. PMID: 20007547 Free PMC article.
-
Combined deficiency of factor V and factor VIII is due to mutations in either LMAN1 or MCFD2.Blood. 2006 Mar 1;107(5):1903-7. doi: 10.1182/blood-2005-09-3620. Epub 2005 Nov 22. Blood. 2006. PMID: 16304051 Free PMC article.
-
Combined deficiency of coagulation factors V and VIII: an update.Semin Thromb Hemost. 2013 Sep;39(6):613-20. doi: 10.1055/s-0033-1349223. Epub 2013 Jul 12. Semin Thromb Hemost. 2013. PMID: 23852824 Free PMC article. Review.
-
Recent developments in the understanding of the combined deficiency of FV and FVIII.Br J Haematol. 2009 Apr;145(1):15-23. doi: 10.1111/j.1365-2141.2008.07559.x. Epub 2009 Jan 16. Br J Haematol. 2009. PMID: 19183188 Free PMC article. Review.
Cited by
-
Clinical, Laboratory, Molecular, and Reproductive Aspects of Combined Deficiency of Factors V and VIII.Semin Thromb Hemost. 2025 Mar;51(2):116-127. doi: 10.1055/s-0044-1789019. Epub 2024 Aug 29. Semin Thromb Hemost. 2025. PMID: 39209292 Free PMC article. Review.
-
Principles of treatment and update of recommendations for the management of haemophilia and congenital bleeding disorders in Italy.Blood Transfus. 2014 Oct;12(4):575-98. doi: 10.2450/2014.0223-14. Blood Transfus. 2014. PMID: 25350962 Free PMC article. No abstract available.
-
The COPII pathway and hematologic disease.Blood. 2012 Jul 5;120(1):31-8. doi: 10.1182/blood-2012-01-292086. Epub 2012 May 14. Blood. 2012. PMID: 22586181 Free PMC article. Review.
-
RNAi targeting LMAN1-MCFD2 complex promotes anticoagulation in mice.J Thromb Thrombolysis. 2024 Dec;57(8):1349-1362. doi: 10.1007/s11239-024-03034-6. Epub 2024 Sep 2. J Thromb Thrombolysis. 2024. PMID: 39222205 Free PMC article.
-
COPII-Dependent ER Export: A Critical Component of Insulin Biogenesis and β-Cell ER Homeostasis.Mol Endocrinol. 2015 Aug;29(8):1156-69. doi: 10.1210/me.2015-1012. Epub 2015 Jun 17. Mol Endocrinol. 2015. PMID: 26083833 Free PMC article.
References
-
- Zhang B, Ginsburg D. Familial multiple coagulation factor deficiencies: new biologic insight from rare genetic bleeding disorders. J Thromb Haemost. 2004;2(9):1564–1572. - PubMed
-
- Nichols WC, Seligsohn U, Zivelin A, et al. Mutations in the ER-Golgi intermediate compartment protein ERGIC-53 cause combined deficiency of coagulation factors V and VIII. Cell. 1998;93(1):61–70. - PubMed
-
- Zhang B, Cunningham MA, Nichols WC, et al. Bleeding due to disruption of a cargo-specific ER-to-Golgi transport complex. Nat Genet. 2003;34(2):220–225. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
