

Axonal α7 nicotinic ACh receptors modulate presynaptic NMDA receptor expression and structural plasticity of glutamatergic presynaptic boutons
- PMID: 20817852
- PMCID: PMC2944730
- DOI: 10.1073/pnas.1007397107
Axonal α7 nicotinic ACh receptors modulate presynaptic NMDA receptor expression and structural plasticity of glutamatergic presynaptic boutons
Abstract
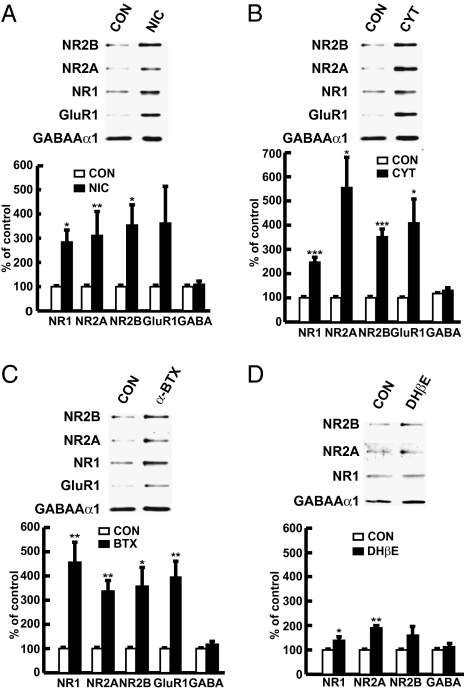
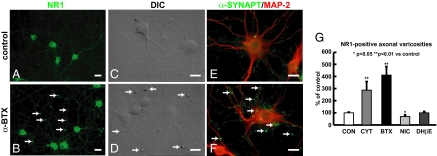
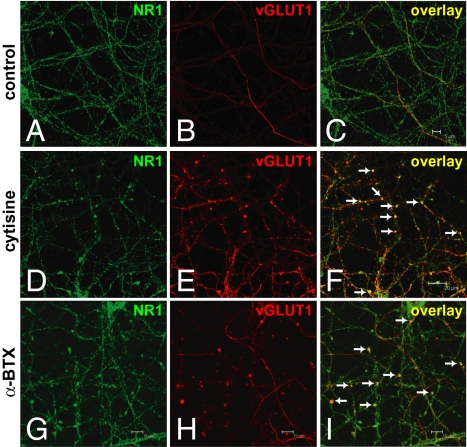
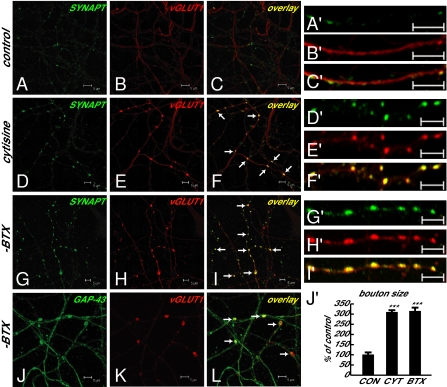
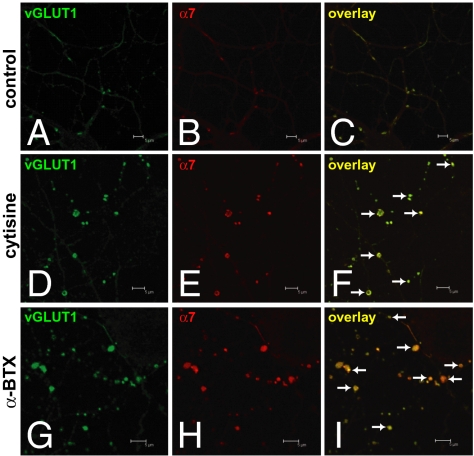
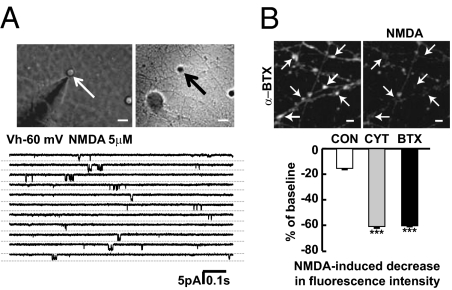
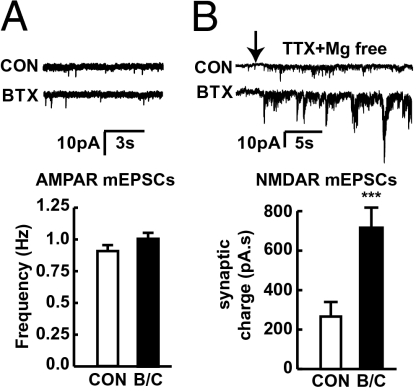
In association with NMDA receptors (NMDARs), neuronal α7 nicotinic ACh receptors (nAChRs) have been implicated in neuronal plasticity as well as neurodevelopmental, neurological, and psychiatric disorders. However, the role of presynaptic NMDARs and their interaction with α7 nAChRs in these physiological and pathophysiological events remains unknown. Here we report that axonal α7 nAChRs modulate presynaptic NMDAR expression and structural plasticity of glutamatergic presynaptic boutons during early synaptic development. Chronic inactivation of α7 nAChRs markedly increased cell surface NMDAR expression as well as the number and size of glutamatergic axonal varicosities in cortical cultures. These boutons contained presynaptic NMDARs and α7 nAChRs, and recordings from outside-out pulled patches of enlarged presynaptic boutons identified functional NMDAR-mediated currents. Multiphoton imaging of presynaptic NMDAR-mediated calcium transients demonstrated significantly larger responses in these enlarged boutons, suggesting enhanced presynaptic NMDAR function that could lead to increased glutamate release. Moreover, whole-cell patch clamp showed a significant increase in synaptic charge mediated by NMDAR miniature EPSCs but no alteration in the frequency of AMPAR miniature EPSCs, suggesting the selective enhancement of postsynaptically silent synapses upon inactivation of α7 nAChRs. Taken together, these findings indicate that axonal α7 nAChRs modulate presynaptic NMDAR expression and presynaptic and postsynaptic maturation of glutamatergic synapses, and implicate presynaptic α7 nAChR/NMDAR interactions in synaptic development and plasticity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Waxman EA, Lynch DR. N-methyl-D-aspartate receptor subtypes: Multiple roles in excitotoxicity and neurological disease. Neuroscientist. 2005;11:37–49. - PubMed
-
- Dingledine R, Borges K, Bowie D, Traynelis SF. The glutamate receptor ion channels. Pharmacol Rev. 1999;51:7–61. - PubMed
-
- Nashmi R, Lester HA. CNS localization of neuronal nicotinic receptors. J Mol Neurosci. 2006;30:181–184. - PubMed
-
- Zhang L, Warren RA. Muscarinic and nicotinic presynaptic modulation of EPSCs in the nucleus accumbens during postnatal development. J Neurophysiol. 2002;88:3315–3330. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
