

SLAM is a microbial sensor that regulates bacterial phagosome functions in macrophages
- PMID: 20818396
- PMCID: PMC3338319
- DOI: 10.1038/ni.1931
SLAM is a microbial sensor that regulates bacterial phagosome functions in macrophages
Abstract
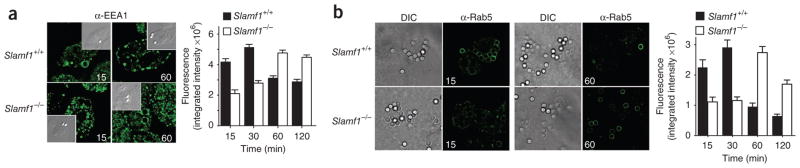
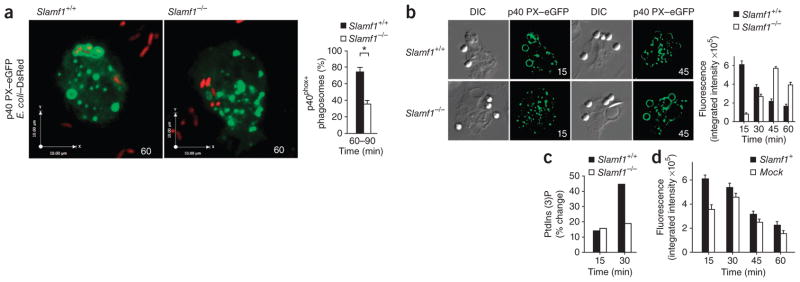
Phagocytosis is a pivotal process by which macrophages eliminate microorganisms after recognition by pathogen sensors. Here we unexpectedly found that the self ligand and cell surface receptor SLAM functioned not only as a costimulatory molecule but also as a microbial sensor that controlled the killing of gram-negative bacteria by macrophages. SLAM regulated activity of the NADPH oxidase NOX2 complex and phagolysosomal maturation after entering the phagosome, following interaction with the bacterial outer membrane proteins OmpC and OmpF. SLAM recruited a complex containing the intracellular class III phosphatidylinositol kinase Vps34, its regulatory protein kinase Vps15 and the autophagy-associated molecule beclin-1 to the phagosome, which was responsible for inducing the accumulation of phosphatidylinositol-3-phosphate, a regulator of both NOX2 function and phagosomal or endosomal fusion. Thus, SLAM connects the gram-negative bacterial phagosome to ubiquitous cellular machinery responsible for the control of bacterial killing.
Conflict of interest statement
The authors declare no competing financial interests.
Figures






References
-
- Stuart LM, Ezekowitz RA. Phagocytosis and comparative innate immunity: learning on the fly. Nat Rev Immunol. 2008;8:131–141. - PubMed
-
- Aderem A, Ulevitch RJ. Toll-like receptors in the induction of the innate immune response. Nature. 2000;406:782–787. - PubMed
-
- Calpe S, et al. The SLAM and SAP gene families control innate and adaptive immune responses. Adv Immunol. 2008;97:177–250. - PubMed
-
- Tatsuo H, Ono N, Tanaka K, Yanagi Y. SLAM (CDw150) is a cellular receptor for measles virus. Nature. 2000;406:893–897. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
