

Upregulation of Tim-3 and PD-1 expression is associated with tumor antigen-specific CD8+ T cell dysfunction in melanoma patients
- PMID: 20819923
- PMCID: PMC2947081
- DOI: 10.1084/jem.20100637
Upregulation of Tim-3 and PD-1 expression is associated with tumor antigen-specific CD8+ T cell dysfunction in melanoma patients
Abstract
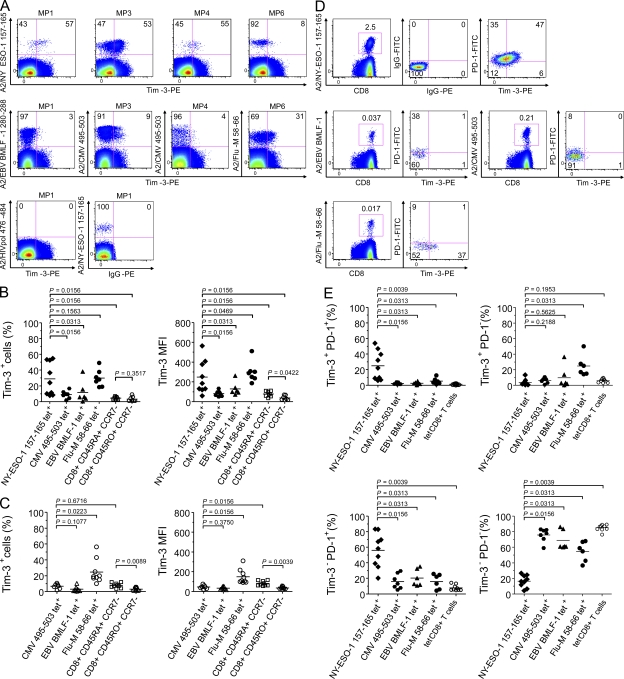
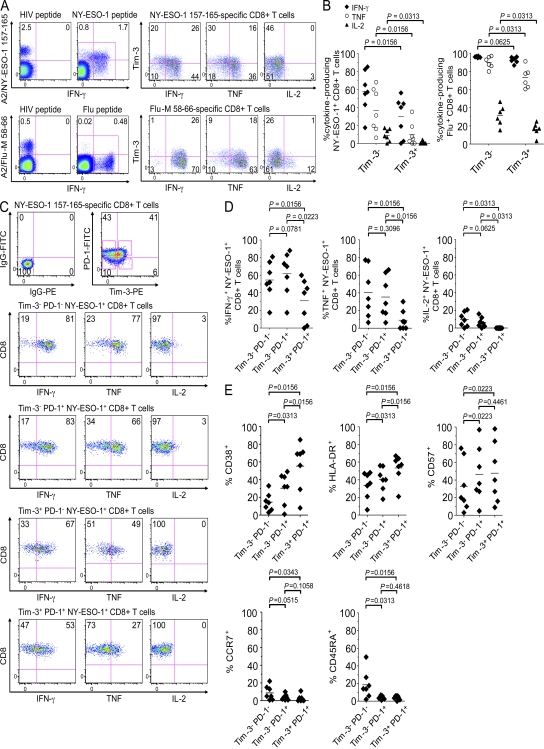
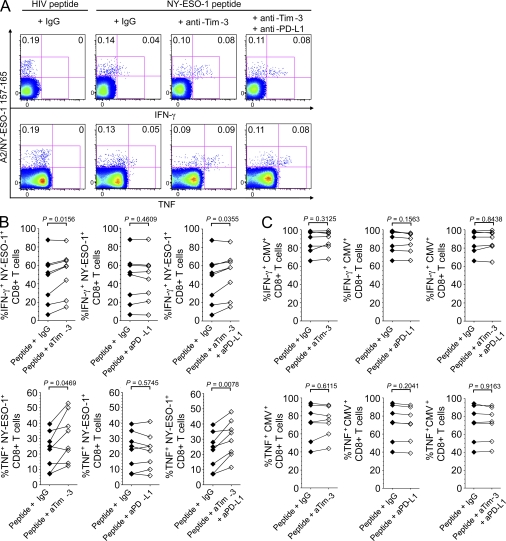
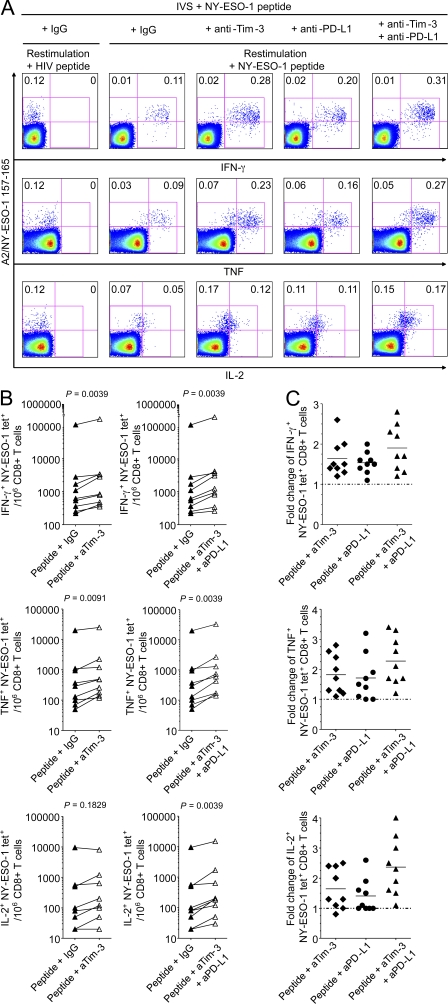
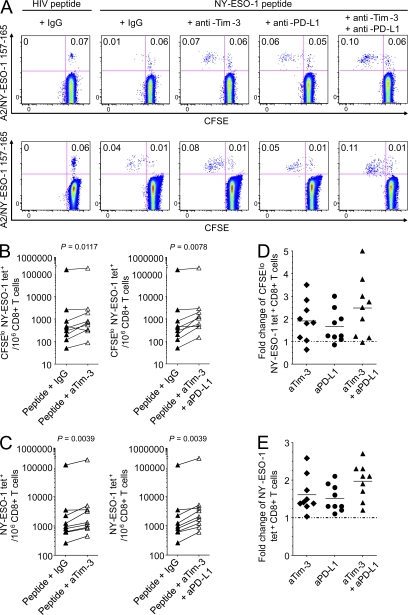
The paradoxical coexistence of spontaneous tumor antigen-specific immune responses with progressive disease in cancer patients furthers the need to dissect the molecular pathways involved in tumor-induced T cell dysfunction. In patients with advanced melanoma, we have previously shown that the cancer-germline antigen NY-ESO-1 stimulates spontaneous NY-ESO-1-specific CD8(+) T cells that up-regulate PD-1 expression. We also observed that PD-1 regulates NY-ESO-1-specific CD8(+) T cell expansion upon chronic antigen stimulation. In the present study, we show that a fraction of PD-1(+) NY-ESO-1-specific CD8(+) T cells in patients with advanced melanoma up-regulates Tim-3 expression and that Tim-3(+)PD-1(+) NY-ESO-1-specific CD8(+) T cells are more dysfunctional than Tim-3(-)PD-1(+) and Tim-3(-)PD-1(-) NY-ESO-1-specific CD8(+) T cells, producing less IFN-γ, TNF, and IL-2. Tim-3-Tim-3L blockade enhanced cytokine production by NY-ESO-1-specific CD8(+) T cells upon short ex vivo stimulation with cognate peptide, thus enhancing their functional capacity. In addition, Tim-3-Tim-3L blockade enhanced cytokine production and proliferation of NY-ESO-1-specific CD8(+) T cells upon prolonged antigen stimulation and acted in synergy with PD-1-PD-L1 blockade. Collectively, our findings support the use of Tim-3-Tim-3L blockade together with PD-1-PD-L1 blockade to reverse tumor-induced T cell exhaustion/dysfunction in patients with advanced melanoma.
Figures
References
-
- Fourcade J., Kudela P., Andrade Filho P.A., Janjic B., Land S.R., Sander C., Krieg A., Donnenberg A., Shen H., Kirkwood J.M., Zarour H.M. 2008. Immunization with analog peptide in combination with CpG and montanide expands tumor antigen-specific CD8+ T cells in melanoma patients. J. Immunother. 31:781–791 10.1097/CJI.0b013e318183af0b - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
